Какие ароморфозы произошли у млекопитающих?
Замечу, что ароморфозы млекопитающих возникли независимо от птиц: это две разные эволюционные ветви. Млекопитающие произошли не от птиц, а от древних пресмыкающихся. Поэтому не удивляйтесь, если некоторые ароморфозы окажутся уже вам знакомы.
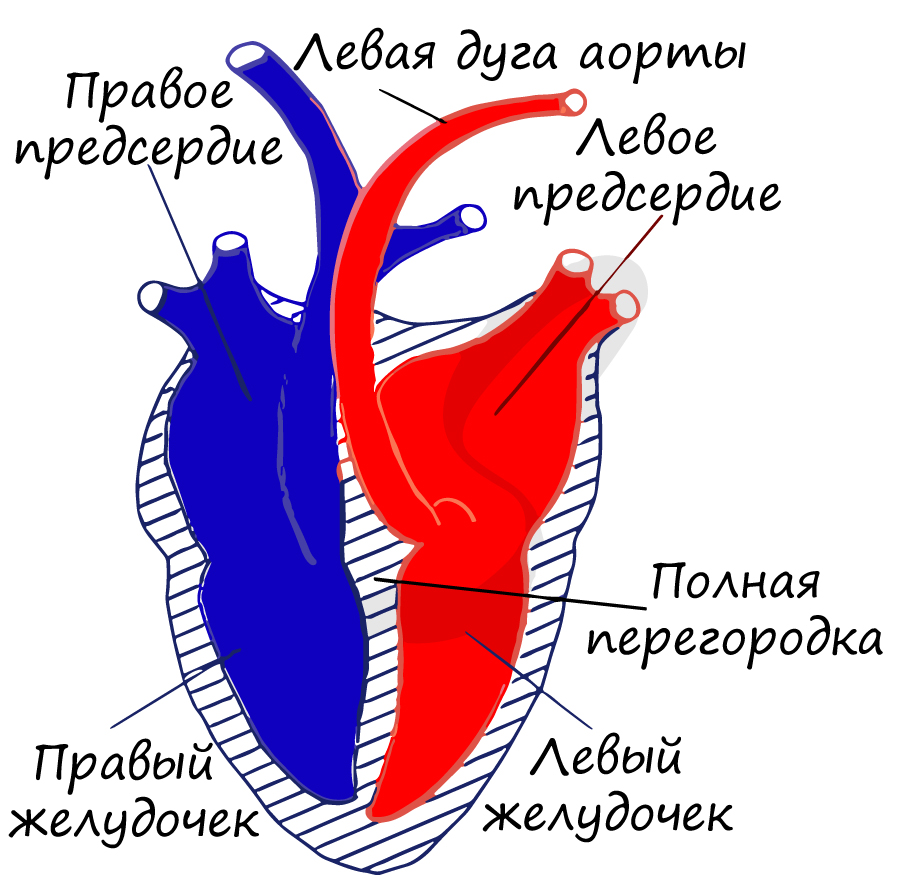
У млекопитающих в сердце возникает полная перегородка: кровь не смешивается, два круга кровообращения полностью отделены друг от друга. Это значительно повышает уровень обмена веществ: температура тела млекопитающих более не зависит от температуры окружающей среды, они приобретают теплокровность (гомотермию).
Развитие коры больших полушарий (КБП), центра высшей нервной деятельности, достигает пика. Такое сильное развитие КБП обеспечивает возникновение сложного поведения, облегчает формирование условных рефлексов, способствуя повышению выживаемости вида.
Для млекопитающих характерна забота о потомстве, проявляющаяся и в уже упомянутом вскармливании молоком, и в том, что детеныши часто живут с матерью несколько лет. Еще во время беременности самка ищет укромное место для рождения потомства, заранее заготавливает корм. Такая забота выглядит удивительно трогательно в диком животном мире.

Волосяной покров помогает более эффективно сохранять тепло, обеспечивая термоизоляцию организму животного. Часть млекопитающих утратила волосяной покров, в связи со вторичноводным образом жизни: они избрали своей средой обитания моря, океаны, где волосяной покров явно был бы лишним и создавал ненужное трение о воду.

По выполняемой функции зубы делятся на резцы, клыки, предкоренные и коренные зубы. У некоторых групп животных, к примеру, у грызунов, резцы приобретают способность к неограниченному росту. Часто хищники имеют хорошо развитые клыки: тигры, львы.

P.S.S. Для вас готов следующий случайный вопрос. Мы сами не знаем, но вас ждет что-то интересное!
© Беллевич Юрий Сергеевич 2018-2021
Текст и опубликованные материалы являются интеллектуальной собственностью Беллевича Юрия Сергеевича. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов вопроса и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Эволюция млекопитающих: описание, ступени, классы
Эволюция — это естественное развитие любых процессов окружающей среды, в число которых входят генетические мутации популяций животных, адаптация, образование новых и вымирание старых видов, изменение отдельных экосистем и, следовательно, всей биосферы в целом.
Маммализация териодонтов
Происхождение и эволюция млекопитающих древнего мира до современных, согласно предположениям исследователей, началась 225 млн лет назад. Это связывают с тем, что некоторые представители животного мира приобрели возможность поднять свой уровень метаболизма, увеличить общую температуру тела и способность самостоятельно ее регулировать. Новые умения сопровождали изменения физического плана:
Появление млекопитающих
 Вам будет интересно: Ганноверская династия: годы, представители, роль в истории Великобритании
Вам будет интересно: Ганноверская династия: годы, представители, роль в истории Великобритании
Поздний меловой период известен тем, что именно в это время появились первые млекопитающие. Древние представители, по сути, это насекомоядные разных видов. Их внешний облик был очень похож: плацентарное теплокровное существо, имеющее серый шерстяной покров и пятипалые конечности. Вытянутый нос имел форму хоботка и помогал животному искать насекомых и личинок.
Большинство ископаемых нашли в меловых отложениях Монголии и Средней Азии. Их предками называют рептилий, входящих в группу синапсидных животных. Именно это группа сформировала подкласс звероподобных существ. Среди них возникли зверозубые представители, которые оказались наиболее близкими к млекопитающим.

Синапсиды
Мезозойская эра создала все условия для благоденствия рептилий со всеми привычными свойствами настоящих ящериц. История запомнила их под названием «динозавры». Зверозубые представители пытались выжить среди них, поэтому вынуждены были сократить размеры тела, уменьшить численность своей популяции и уйти в тень, заняв второстепенную природную нишу, отдав главенство другим животным. Их расцвет начнется позже в результате климатических изменений и последовавшим за этим вымиранием ящеров.
Дииктодон
 Вам будет интересно: Южная Бессарабия: география, политика, управление. Полоса Кагул-Измаил-Болград
Вам будет интересно: Южная Бессарабия: география, политика, управление. Полоса Кагул-Измаил-Болград
Возраст найденных останков — от 252 млн лет. Это одно из самых древних животных, у которого были бивни на нижней челюсти. Длина его тела не превышала 80 сантиметров. Дииктодон жил на территории современной Европы еще до появления первых динозавров. Значительно позже именно от него произошли предки млекопитающих.
Двиния
Это животное было около 50 сантиметров в длину. Оно имело шерстяной покров и зубы, схожие по строению с челюстным аппаратом млекопитающих. Отличительные черты:
Скорее всего, двиния была всеядной. Несмотря на много сходных черт, ее мозг был более примитивным, чем у простейших млекопитающих.
Плацериас
Возраст найденных останков — от 215 млн лет назад. Их относят к группе терапсид, от которой впоследствии также произошли млекопитающие.
Дидельфодон

Возраст останков — от 65 млн лет назад. Возможная территория проживания — США, Монтана, Австралия, Южная Америка. Это одно из древних сумчатых животных, от которых впоследствии произошли опоссумы.
Длина дидельфодона не превышала 1 метра, а вес был около 20 килограмм. Он имел острое зрение, поэтому есть предположение, что зверь был ночным обитателем. Питался мелкими животными, насекомыми, яйцами динозавров и любой найденной падалью.
Кондилартр
Протитан
 Вам будет интересно: Советские солдаты в Афганистане: статистика, форма, фото
Вам будет интересно: Советские солдаты в Афганистане: статистика, форма, фото
Большинство останков найдено в Северной Америке. Их возраст определяют на уровне 35 млн лет назад. Согласно предположениям исследователей, их образ жизни напоминал современных бегемотов. Дневное время они лежали в воде на мелководье, а вечером шли на берег за травой.
Австралопитек

Это большая человекообразная обезьяна. Считается, что его родственники стали непосредственными предками современных людей. Время их появления приходится на период от 6 млн лет назад.
Они жили на территории Африки небольшими группами, в состав которых входили 2 или 3 самца, несколько самок и общее потомство. Основу их рациона составляли растения и семена. Именно это стало причиной уменьшения клыков и началом прямохождения, так как среди высоких зарослей, передвигаясь на четырех ногах, было трудно увидеть хищника.Эволюция мозга млекопитающих была пока еще в начальной стадии, поэтому объем серого вещества уступал содержимому черепной коробки древних людей.
Африканский австралопитек — примат, чей рост не превышает 150 сантиметров. Исследователи предполагают, что он ловко пользовался камнями, ветками и обломками костей, облегчая свой труд. Его линия берет свое начало от афарского австралопитека, который считается предком человеческого рода.
Неандерталец

Поздний представитель человеческого рода. Считается, что неандертальцы появились на территории Африки 400 тыс. лет назад. Впоследствии они расселились по Европе и Азии (в течение ледникового периода). Последние члены популяции вымерли 40 тыс. лет назад.
Очень долго все исследователи видели в неандертальце единственного предка современных людей. Сейчас популярна теория, что оба вида (неандертальцы и современные люди) берут свое начало от одного предка. Определенный промежуток времени они существовали по соседству.
Рост обычного неандертальца составлял около 163 сантиметров, телосложение было крепким и мускулистым, приспособленным к территориям с тяжелыми условиями проживания. Его череп был вытянутой формы, с крепкими и сильными челюстями, ярко выраженными надбровными дугами. Строение черепной коробки указывает на острое зрение и примитивную речь. Они умели пользоваться простыми орудиями труда и выработали своеобразный социум.
Ранние млекопитающие
У древних представителей потовые железы видоизменились, образовав млечные. Вероятно, сначала они не кормили свое потомство, а выпаивали, обеспечивая их постоянным доступом к жизненно необходимой жидкости и соли. Следующими изменились зубы, разделив первых млекопитающих на две группы — кюнеотериид и морганукодонтид.
Другая линия, под названием пантотерии, лучше приспособилась к стремительно меняющимся условиям жизни. Внешне они напоминали маленьких зверят, которые питаются насекомыми, яйцами и потомством других животных. На этот промежуток времени размер их мозга был слишком мал, но уже больше, чем у других зверозубых представителей. Конец мезозойской эры оказался решающим для этого вида, разделив его на две самостоятельные разновидности — высшие плацентарные и низшие сумчатые.
В начале мелового периода появились плацентарные животные. Как показала дальнейшая эволюция млекопитающих, этот вид оказался довольно успешным.

Развитие древних млекопитающих до современных животных
Зверозубые существовали до периода верхнего Триаса. Окаменелые останки древних млекопитающих находят в юрских отложениях.
В дальнейшем, от бугорчатозубых животных произошли плацентарные и сумчатые млекопитающие. В начале меловой эры плацентарные разделились, образовав линии китообразных и грызунов. Те из них, которые питались насекомыми, сформировали множество линий: летучий мышей, приматов, неполнозубых и так далее. Хищно копытная разновидность отделилась, образовав самостоятельный биологический вид, который со временем дал начало хищным и копытным животным. От древнейших хищных, так называемых креодонтов, произошли ластоногие, от первых копытных — парнокопытные, непарнокопытные и хоботные. В конце кайнозойской эры плацентарные млекопитающие заняли главную природную нишу. Из них сформировался 31 отряд животных, 17 из которых живут и сегодня.
Наиболее древними млекопитающими считаются те, которые питались насекомыми. Внешне они напоминали маленьких зверят, способных жить на земле и деревьях. Насекомоядные, передвигающиеся по деревьям, в процессе эволюции конечностей млекопитающих, начали планировать, а чуть позже и летать, образовав отряд рукокрылых. Наземные формы увеличивались в размерах, что позволило им перейти на охоту за более крупной дичью, что позволило им образовать класс креодонтов. Со временем они уступили место предкам современных животных из отряда Garnivora. В Неогене появились известные на весь мир саблезубые кошки.
На протяжении Палеогена хищники сформировали две параллельные линии: ластоногие и наземные хищные млекопитающие. Ластоногие заняли все водоемы, и стали морскими царями.

Отдельные представители креодонтов, полностью поменявшие свой привычный рацион на растительную пищу, стали предками кондилартров, то есть первых копытных.
С началом Эоцена от насекомоядных отделились и сформировали самостоятельные биологические виды предки грызунов, трубкозубых, приматов и неполнозубых.
 Вам будет интересно: Дмитрий Иоаннович, сын Ивана Грозного: дата рождения, краткая биография, причина смерти и канонизация
Вам будет интересно: Дмитрий Иоаннович, сын Ивана Грозного: дата рождения, краткая биография, причина смерти и канонизация
Эволюция птиц и млекопитающих продолжалась на протяжении всего кайнозойского периода. Появились первые цветы, ставшие неотъемлемым элементом ежедневного рациона млекопитающих. Периодически менялась экология, вынуждая животных приспосабливаться к новым условиям жизни. Древние птицы и млекопитающие достигли в эволюции поставленных целей и постепенно исчезали, а их потомство с каждым новым поколением становилось более развитым и совершенным. Но процесс разделения континентов сформировал отдельные области, изолированные от остального мира, в которых очень долго проживали первоначальные формы животных.
В период расцвета сумчатых от других материков отделилась Австралия. Со временем Южная Америка отошла от Северной. В результате этого биологические виды, проживающие на этой территории, развивались самостоятельно.
Главная природная ниша на территории Южной Америки осталась за сумчатыми, которые ввиду отсутствия конкуренции продолжили свое развитие. Из маленьких плотоядных существ, не превышающих своими параметрами размеры опоссума, они превратились в огромных животных, известных под названием саблезубых тигров.
В процессе эволюции класса млекопитающих появлялись гигантские формы муравьедов, броненосцев и ленивцев. Стабильное сосуществование сумчатых и плацентарных млекопитающих закончилось в конце Плиоцена. В это время образовался перешеек, соединивший Северную и Южную Америку. Впервые за очень долгий промежуток времени животные южной части встретились с северными соседями. Последние были наиболее развитыми, поэтому без труда истребили сумчатых и копытных. Лишь гигантские броненосцы и ленивцы смогли пройти дальше северной области, достигнув территории Аляски.
На территории Евразии и Северной Америки проходили все ступени эволюции млекопитающих копытные животные и слоны. Благодаря палеонтологам детальнее разобрано развитие лошадей, которое в основном происходило на территории Северной Америки. Их предком считают гиракотерию или эогиппуса, чье существование приходится на период Палеоцена. Рацион гиракотерия составляла жесткая листва кустарников, а их передвижение в окружающем пространстве было очень быстрым.
Древние пастбища дали возможность лошадям не искать себе пищу, общипывая кусты и молодые побеги, а спокойно пастись на обширных равнинах. Некоторые представители вида остались бродить в широких кустарниках, сохранив размеры пони. Они сформировали гиппарионовую фауну, со временем распространившуюся по территориям Евразии и Северной Америки. Основу их рациона составляли молодые растения и листья на деревьях и кустарниках. У них была конкуренция в лице маленьких носорогов с длинными конечностями, чьи особи не выдержали натиска лошадей и вымерли.
Остальные носороги выглядели, как нынешние гиппопотамы. Были такие виды, которые выросли до впечатляющих размеров. Самым известным из них стал белуджитерий — крупнейшее млекопитающее, из когда-либо существовавших на Земле. Рост отдельных представителей вида превышал 6 метров, что позволяло им доставать до листьев и побегов самых высоких деревьев.
Развитие слонов было не менее сложным. Их окончательное становление произошло в период Неогена. В это время кайнозойские формы слоновьих предков стали иначе пережевывать пищу — вперед и назад, передвигаясь в одном направлении. Именно резкое изменение жевательного аппарата спровоцировало формирование всемирно известных особенностей слоновьей головы.
Меловой период стал переломным и для отряда приматов. Они появились 80 млн лет назад, и своим видом напоминали современных животных, например, долгопята или лемура. С началом палеогена началось их разделение на низших и человекообразных представителей. Около 12 млн лет назад появился рамапитек — первый примат, имеющий внешнее сходство с человеком. К местам его обитания относят Индию и Африку.
5 млн лет назад в Африке появились первые австралопитеки — близкие родственники расы, которые еще относятся к виду приматов, но умеют ходить на двух ногах и ежедневно применять самодельные орудия. Около 2500000 лет назад они начали переходить на человеческий труд, что доказывают уникальные останки австралопитеков, найденные палеонтологами на территории Восточной Африки. Начало Палеолита оставило свой след в истории тем, что в этот период появились первые люди.
Главные черты царей мира животных
Благодаря эволюции, млекопитающие добились высшего класса позвоночных, которые заняли главную ступеньку в животном царстве. Их общая организация достойна отдельного внимания:
Такие качества обеспечили распространение млекопитающих на земле, в воде и воздухе. Их царствование не дошло лишь до Антарктического континента. Но даже там можно встретить отголоски этой власти в лице китов и тюленей.
В тени динозавров. Родословная млекопитающих
Млекопитающие – уникальная группа живых организмов в истории нашей планеты, представители которой освоили передачу значительной часть информации от поколения к поколению негенетическим путем. С момента происхождения и развития жизни эта эволюционная тенденция наблюдалась постоянно в разных эволюционных линиях живых существ. Но только в стволе млекопитающих она достигла своего наиболее яркого и полного выражения, породив в итоге «феномен человека». Именно это привело к формированию ноосферы, это определило и определяет в настоящее время структуру биосферы. Поэтому так важно понять роль и место млекопитающих в истории развития живого.
Первые млекопитающие появились на нашей планете в позднем триасе, примерно 220 млн лет назад. Это не так уж много, если учесть, что первые микроорганизмы появились более 3,5 млрд лет назад, многоклеточные – примерно 1,5 млрд лет назад, а первые позвоночные – в раннем ордовике, т. е. около 500 млн лет назад.
Эти две сотни миллионов лет были полны драматизма. В основе сюжета исторической драмы лежало противостояние двух разных эволюционных групп, оспаривавших друг у друга господствующее положение. Обе группы произошли в позднем палеозое от лабиринтодонтов – древнейших амфибий, унаследовавших от своих прямых предков – кистеперых рыб – плоский, напоминающий панцирь череп с отверстиями лишь для глаз и ноздрей.
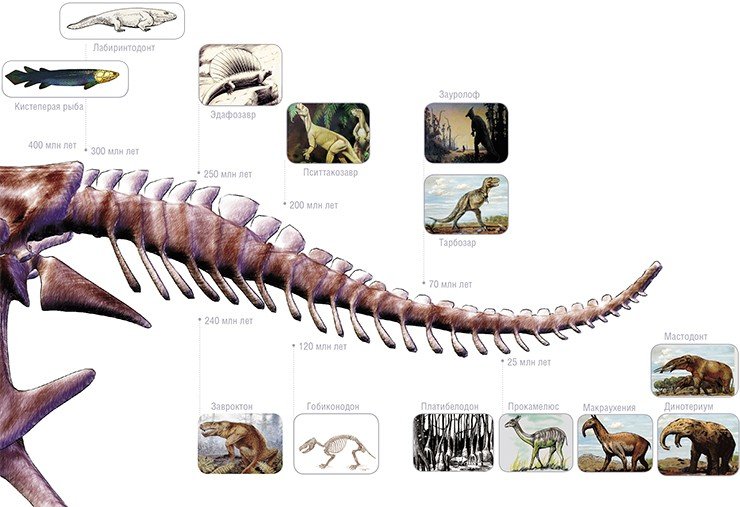
Примитивные легкие и отсутствие подвижных ребер – «мехов» для нагнетания воздуха – заставляло амфибий использовать ротовую полость как дополнительный орган дыхания (так же, как дышит лягушка), в результате чего голова у них достигала гигантских размеров – до трети длины тела.
Такой череп тормозил развитие жевательной мускулатуры и челюстного аппарата, да и дальнейшую эволюцию группы в целом. Появление более совершенной дыхательной системы позволило снять с ротовой полости эту дополнительную «нагрузку», и череп начал быстро совершенствоваться. Его поперечный профиль из уплощенного становится сводчатым, что резко увеличивает прочность конструкции. В крыше черепа появляются отверстия для выхода челюстной мускулатуры.
Подобная модернизация черепа протекала двумя различными путями: в группе так называемых диапсид появилось два таких отверстия, у синапсид – одно. В этом и состоит внешнее анатомическое различие двух самых крупных групп наземных позвоночных, давших мощную радиацию в позднем палеозое и мезозое. Но их исторические судьбы оказались совершенно различными.
Динозавры первыми «встали на ноги»
Одно из основных направлений эволюции диапсид – формирование ажурной и подвижной структуры черепа.
Кроме того, у диапсид могут постоянно отрастать зубы взамен поломанных или утраченных. За это «удобство» приходится дорого платить: беспорядочная смена зубов исключает окклюзию (точную подгонку верхних и нижних зубов друг к другу). Поэтому у диапсид отсутствует зубной аппарат режущего типа.
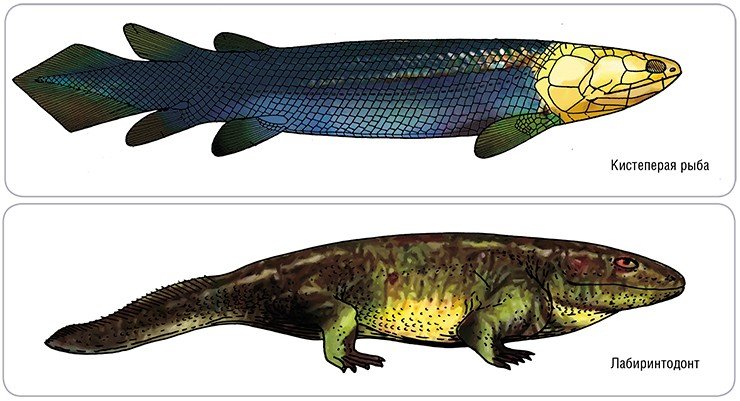
Диапсиды по-своему решили общую для наземных позвоночных проблему передвижения на суше. Многие из них перешли к бипедии, т. е. к хождению на двух ногах, что сопровождалось редукцией передних конечностей, а некоторые вовсе утратили ноги, как, например, змеи.
Ко всему, диапсиды, по-видимому, никогда не обладали гомойотермией – физиологически обусловленной стабильной температурой тела. Исключение составляют только птицы, да у некоторых крупных динозавров инерционное сохранение более или менее постоянной температуры обеспечивалось благодаря большим размерам тела.
К ДИАПСИДАМ (т. е. имеющим в черепе два отверстия для выхода челюстной мускулатуры и две скуловые дуги) относятся динозавры в широком смысле, их триасовые предшественники – архозавры и потомки: крокодилы, ящерицы, змеи, а также птицы.
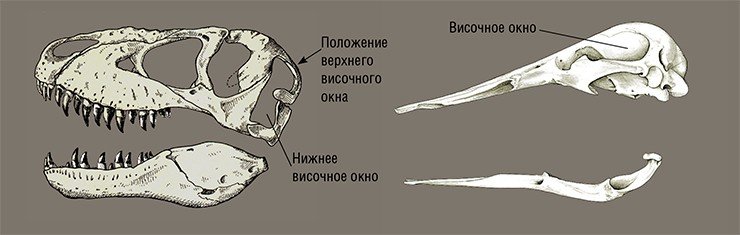
Для диапсид характерна ажурная и подвижная структура черепа. Например, ящерицы и змеи, раскрывая рот, не только опускают вниз нижнюю челюсть, но могут также приподнимать и опускать переднюю часть верхней челюсти. Нижнечелюстной сустав может раздвигаться. Обе половинки нижней челюсти соединены между собой эластичными связками. У диапсид фактически нет мозговой коробки: полость, в которой расположен мозг, открыта спереди и представляет собой подобие трубки.
Сам мозг как у древних, так и у современных диапсид (кроме птиц) очень примитивен.
Конструктивное единство элементов черепа у синапсид позволило им уже в триасе сформировать монолитную черепную коробку. Число костей нижней челюсти у синапсид неуклонно уменьшалось, и в итоге из шести костей осталась только одна – зубная.
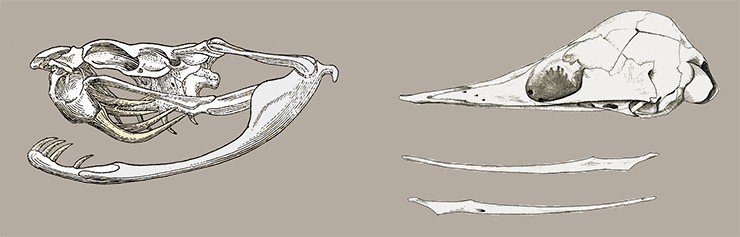
Такое монолитное строение черепа стало, по существу, преадаптацией к дальнейшему развитию мозга.
Млекопитающие (а также пеликозавры и терапсиды, в том числе териодонты — «зверозубые рептилии») относятся к СИНАПСИДАМ, у которых в черепе имеется только одно отверстие (височное окно) для выхода челюстной мускулатуры
В отличие от диапсид, у синапсид никогда не формировалась подвижная конструкция черепа: со временем последний, наоборот, становился все более монолитным. Это можно рассматривать как преадаптацию к совершенствованию мозга, которое, впрочем, началось гораздо позже.
Со временем количество зубов и их положение в челюсти у синапсид стабилизировалось, что сделало возможным окклюзию. Это, в свою очередь, позволило осуществлять первичную обработку пищевого комка, чего никто из диапсид – ни динозавры, ни современные крокодилы, ящерицы и змеи – делать не способны.
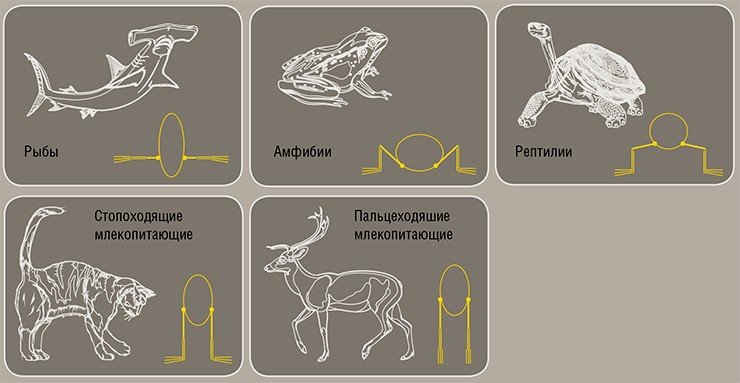
В отношении опорно-двигательного аппарата синапсиды вначале уступали диапсидам: ни в перми, ни в триасе они так и не не научились передвигаться на двух ногах. Передвижение же на четырех конечностях эффективно, только если существо имеет вертикально поставленные конечности, а пермо-триасовые синапсиды как бы «висели» между своими ногами. Латеральное (боковое) положение конечностей сохраняли еще и некоторые мезозойские млекопитающие – такое положение крайне неэкономно с точки зрения расхода мышечной энергии
В совершенствовании локомоторного аппарата синапсиды вначале уступали диапсидам. Перевести ноги в вертикальное положение синапсидам удалось лишь в конце триаса, уже на стадии млекопитающих – для этого потребовались очень крупные преобразования опорно-двигательного аппарата.
Что касается гомойотермии, то, в отличие от диапсид, первые попытки перехода к ней, по-видимому, были у синапсид уже в карбоне. Вспомним пеликазавров с их огромным «парусом» на спине, который специалисты все более склонны рассматривать как средство стабилизации температуры тела.
Диапсиды и синапсиды: великое противостояние
Именно синапсиды первыми в истории Земли создали в пермском периоде богатую и разнообразную фауну позвоночных суши. Только два региона сохранили для нас в большом количестве ее окаменевшие остатки: Южная Африка и Русская платформа (бассейн Северной Двины и Башкирия). Среди пермских синапсид были крупные сильные хищники размером с тигра, с огромными кинжаловидными клыками – горгонопсиды (иностранцевия и др.); были охотники на мелкую добычу, аналоги куньих и псовых; существовали и растительнояды, например, листрозавры, каннемейериды – большие тяжеловесные животные.

К началу юры диапсиды «переиграли» синапсид, очевидно, в основном именно благодаря изобретению двуногой локомоции. Хищные диапсиды (динозавры) полностью вытеснили хищных синапсид. Интересно, что растительноядные синапсиды пережили свое время и сосуществовали с хищниками из диапсид. Как будет рассмотрено ниже, растительноядность вообще, в том числе на базе диапсид, формировалась в процессе эволюции «с трудом». В целом же биота синапсид была почти нацело «стерта» сообществом диапсид, господство которых продолжалось с конца триаса до конца мела.

Расцвет диапсид закончился в позднем мелу, однако многие диапсиды (крокодилы, змеи и ящерицы) прекрасно существуют и в наши дни, а птицы делят с млекопитающими господствующее положение среди позвоночных животных суши. Но история синапсид на этом не закончилась, напротив – в кайнозое для них наступила новая эпоха расцвета. Как же синапсиды пережили долгий период господства диапсид, какие изменения они претерпели за это время, и чем было обусловлено их «возрождение» уже в виде млекопитающих?
Давид против Голиафа
Первые представители млекопитающих произошли в позднем триасе от мелких, эволюционно продвинутых териодонтов. Эти неспециализированные плотояды сумели закрепиться в динозавровой биоте лишь потому, что были очень мелкими, размером с мышь, что позволило им выйти из-под пресса хищных динозавров.

Например, для тиранозавра высотой до 5 м и весом 1,5—2 т охота на жертву, вес которой 10—20 г, была не только бессмысленной, с точки зрения энергетических затрат, но и безуспешной. Это поставило мезозойских синапсид вне пищевых цепей и блоков динозавровой биоты и дало им возможность просуществовать «в тени» динозавров более 150 млн лет, что составляет примерно две трети истории класса млекопитающих.
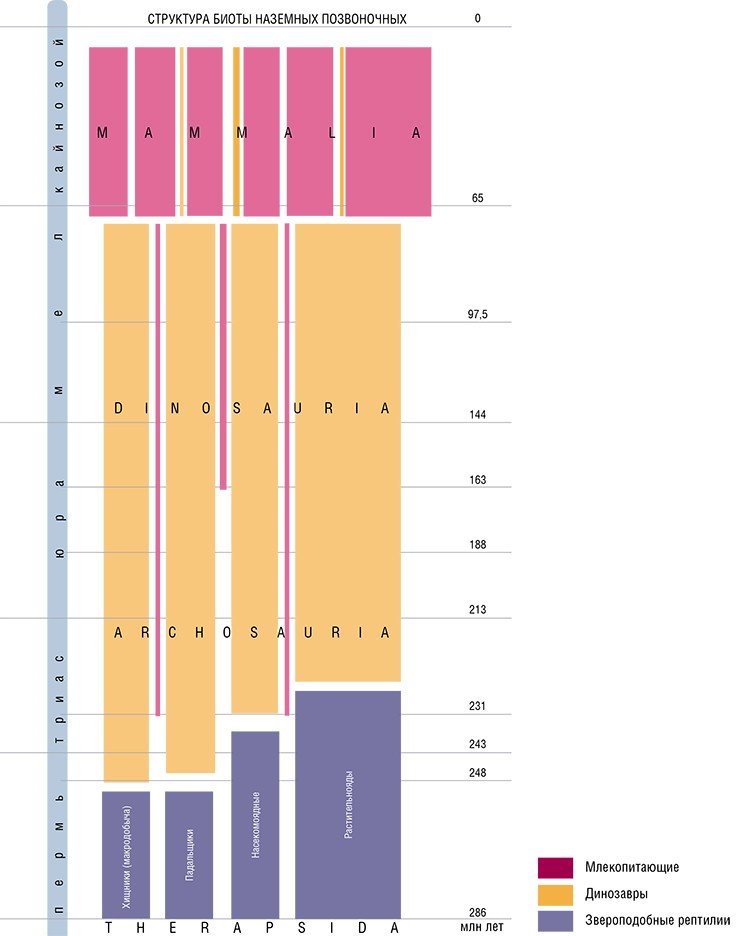
Две трети своей истории млекопитающие буквально «проскакивали между пальцами» крупных хищных динозавров благодаря своим мелким размерам
Важным событием в эволюции наземной биоты было появление многобугорчатых в юрском периоде: фактически впервые в истории Земли млекопитающие сформировали эффективного растительнояда в мелком размерном классе. Дело в том, что переваривать грубую растительную пищу позвоночные способны только с помощью симбиотических кишечных микроорганизмов. Все растительноядные диапсиды были очень крупными, потому что большая масса тела обеспечивала инерционное сохранение относительно стабильной температуры, необходимой для существования кишечной флоры. В мелком размерном классе такое невозможно: кишечная флора будет погибать при любом охлаждении тела (например, ночью). Поэтому эффективное растительноядение у мелких позвоночных возможно только при наличии гомойотермии, присущей млекопитающим.

Первые относительно крупные теплокровные хищники-млекопитающие размером с кошку представляли ощутимую угрозу для детенышей динозавров мелового периода
Остальные млекопитающие юры и мела были также мелкие существа, преимущественно плотоядные. Исключение составляют триконодонты. В юре и в раннем мелу они, как и все другие млекопитающие, были очень маленькими, размером не больше мыши. Однако во второй половине мела среди них появился макрохищник, гигант среди млекопитающих, достигавший размеров современной кошки или вирджинского опоссума (Jenkins, Schaff, 1988).
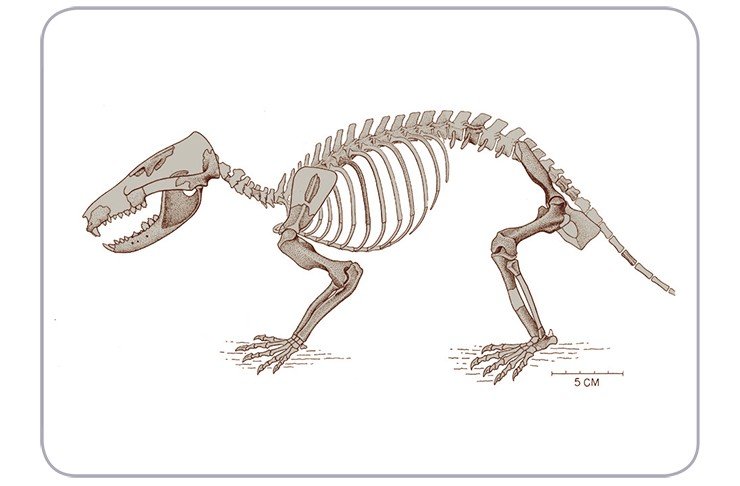
До этого момента биоты млекопитающих и динозавров практически не перекрывались, но теперь ситуация изменилась. Детеныши динозавров, только что вылупившиеся из яиц, были не крупнее курицы, имели хрящевой скелет, а по ночам не могли активно двигаться из-за охлаждения. Наверняка, они были легкой и желанной добычей триконодонтов. Эти агрессивные хищники, способные охотиться по ночам благодаря своей теплокровности, могли существенно повлиять на судьбу выводков меловых динозавров (Агаджанян, 2003). Эта гипотеза недавно получила подтверждение: китайские палеонтологи описали триконодонта Repenomamus giganticus с крупными островершинными зубами и массивной челюстью (длина его тела превышала 1 м, а вес достигал 12—14 кг), в желудке которого были найдены остатки молодого растительноядного динозавра (Hu et al., 2005).
Роль мезозойских млекопитающих в структуре биоты Земли не ограничивалась подобным ощутимым прессом на динозавров. У вышеупомянутых многобугорчатых щечные зубы имели структуру коронки по типу хомякообразных, а резцы по форме и строению напоминали резцы грызунов. С точки зрения функции и положения в сообществе многобугорчатые являлись высокоспециализированными «грызунами». Анализ их жевательного аппарата показал, что они питались довольно крупными объектами (Gambaryan, Kielan-Jaworowska, 1995). В юре и первой половине мела это могли быть орешки голосемянных (цикадовых, гинговых, араукариевых), в течение второй половины мела и в кайнозое – плоды покрытосемянных, в том числе бука, дуба и др.
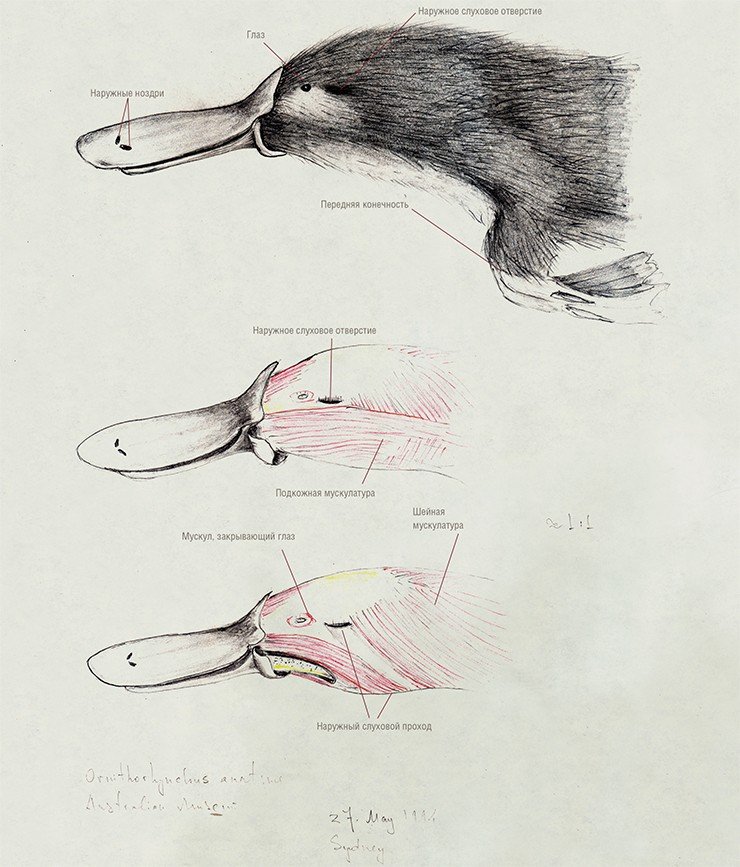
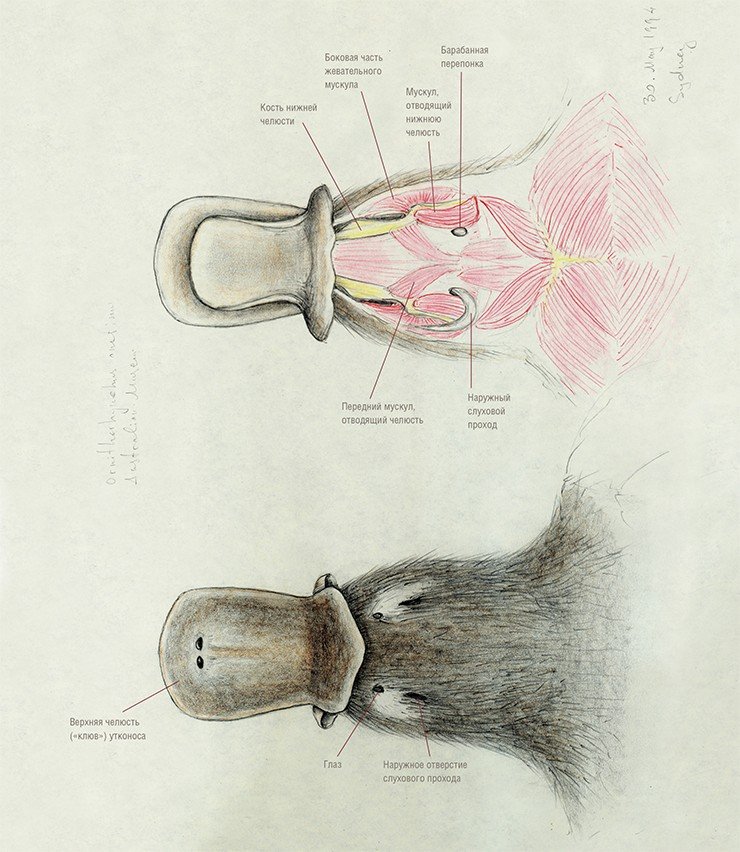
Радиации многобугорчатых и покрытосемянных в начале второй половины мела совпадают по времени и, по-видимому, были взаимообусловлены, т. е. эти млекопитающие могли играть важную роль в расселении голосемянных, а затем и покрытосемянных. Адаптивный тип этой группы млекопитающих был настолько эффективен, что многобугорчатые перешагнули рубеж мел—кайнозой и просуществовали достаточно долго, до конца олигоцена, успешно конкурируя одно время даже с грызунами.
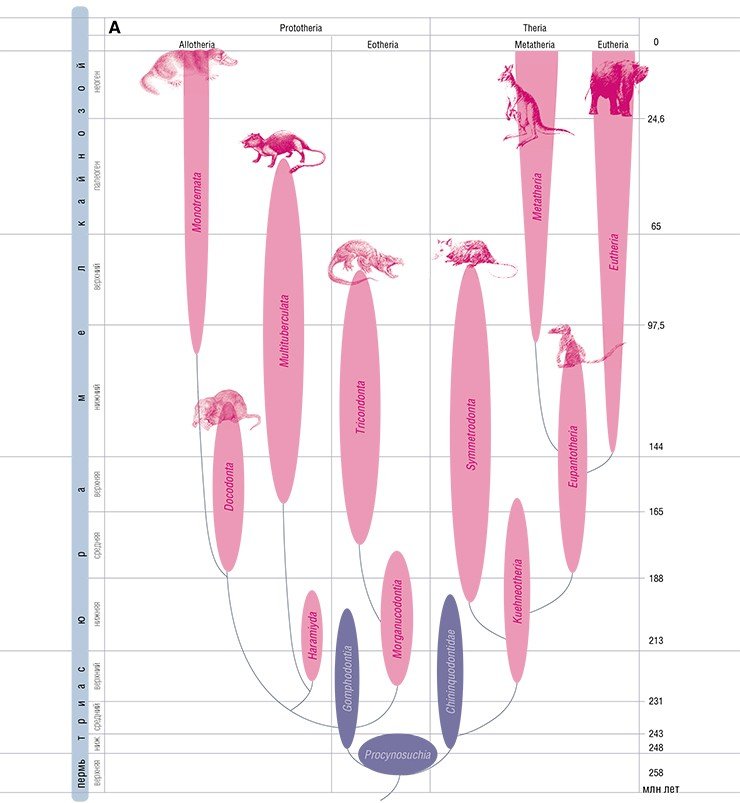
Современные млекопитающие подразделяются на два подкласса: прототерии и терии. И те и другие были известны уже в позднем триасе. В мезозойских сообществах млекопитающих господствовали прототерии, а терии были «изгоями». В кайнозое ситуация изменилась на противоположную. Чтобы понять причины этого, необходимо сделать экскурс в анатомию.
Между молоточком и наковальней
Имеющиеся данные по строению черепа (Kermack, Kielan-Jaworowska, 1971) и среднего уха однопроходных подтвердили глубокие различия между прототериями и териями, что дает основание предполагать, что они произошли от разных групп триасовых териодонтов. Напомним, что в ходе эволюции нижняя челюсть синапсид постепенно теряла кости, расположенные сзади от зубной кости, которая в свою очередь неуклонно увеличивалась, разрастаясь назад. На стадии первых млекопитающих она пришла в соприкосновение с чешуйчатой костью черепной коробки. В результате возникло новое сочленение нижней челюсти с черепом.
Судя по анатомическим особенностям, разные эволюционные линии млекопитающих разошлись еще на уровне предков
У млекопитающих позднего триаса функционировали оба сочленения: старое (сочленованная кость нижней челюсти – квадратная кость черепной коробки) и новое (зубная кость – чешуйчатая кость). Затем старый сустав исчез, а новый сохранился.
Кости, формировавшие старый сустав, преобразовались в элементы звукопередаточного аппарата. Угловая и сочленованная кости нижней челюсти превратились в барабанную кость и молоточек, квадратная кость черепной коробки – в наковальню (еще один элемент звукопередающего механизма, стремечко, имелся уже у рептилий).
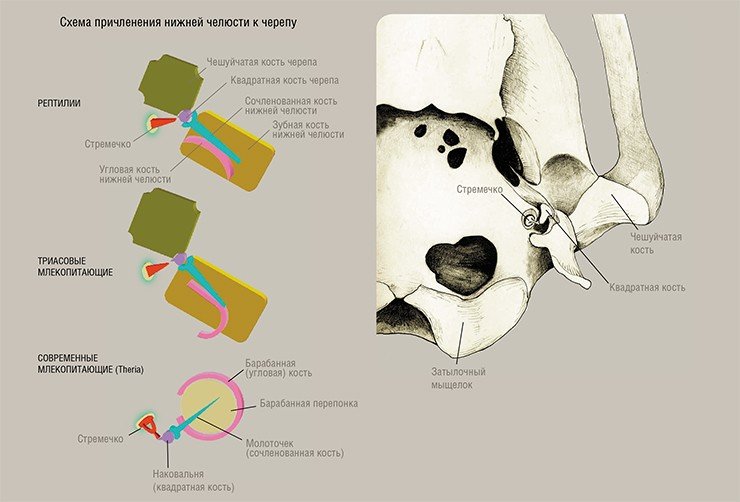
Строение и эволюционная история барабанной кости, наковальни и молоточка различна у прототериев и териев.
По-видимому, именно эти особенности определили различия при модернизации черепа у этих двух групп млекопитающих. Черепа у всех прототериев очень однотипные и практически не менялись на протяжении 200 млн лет вплоть до настоящего времени. У териев, напротив, трехчленность звукопередающего механизма сделала возможной почти неограниченную модернизацию черепа в процессе эволюции и породила многообразие их строения.
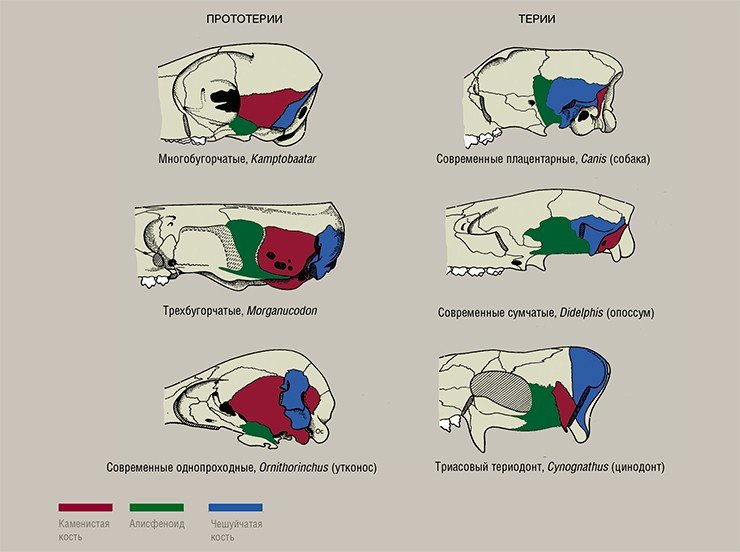
Правота этих рассуждений подтверждается тем, что начиная примерно с олигоцена, когда у териев завершилось формирование основных типов черепов, у многих из них срастаются молоточек и наковальня (Weber, 1927). Когда отпадает нужда хотя бы в одном сочленении, эволюция от него избавляется.
Мезозойские прототерии и терии различались также по строению зубного аппарата. По-видимому, изначально высокая специализация зубов прототериев также препятствовала их дальнейшей эволюции: будучи высоко адаптированными к своей динозавровой биоте, они «заблокировали» возможность изменения зубного аппарата.
Все эти принципиальные различия позволяют настаивать на том, что эволюционные линии прототериев и териев разошлись еще на уровне их предков – териодонтов. Современные однопроходные – это, по существу, животные териодонтового уровня, которые в условиях длительной изоляции на Австралийском континенте сохранили до наших дней многие признаки триасовых предков млекопитающих.
Колыбель для мозга
Что же касается обеих современных групп териев – сумчатых и плацентарных – то их представители известны начиная с раннего мела. Однако совершенная плацентарность, проявляющаяся в длительном вынашивании детенышей, по-видимому, развилась не раньше конца мела или начала кайнозоя. Об этом свидетельствуют так называемые сумчатые кости, имеющиеся у териодонтов, всех современных однопроходных и сумчатых.
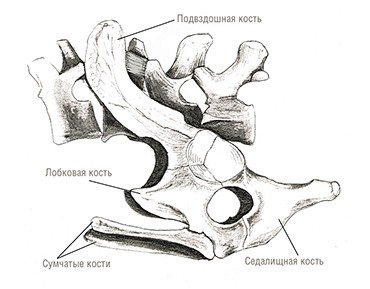 Эти кости имелись и у ранних плацентарных вплоть до конца мела (Novacek et al., 1997). Однако с начала кайнозоя судьбы сумчатых и плацентарных разошлись: они оказались разделенными на двух блоках континентов (Северном и Южном), и их развитие пошло различными путями.
Эти кости имелись и у ранних плацентарных вплоть до конца мела (Novacek et al., 1997). Однако с начала кайнозоя судьбы сумчатых и плацентарных разошлись: они оказались разделенными на двух блоках континентов (Северном и Южном), и их развитие пошло различными путями.
До возникновения совершенной плацентарности уровень развития головного мозга у всех млекопитающих был почти одинаков. Сумчатые, обитатели южных материков, сохраняют примитивное строение мозга и по сей день. Почему так произошло? В целом сумчатость, как тип размножения, выгоднее для сохранения популяции, чем плацентарность. Самка сумчатых в случае опасности, в отличие от плацентарных, просто выбрасывает детеныша из сумки и убегает. Новое зачатие у сумчатых возможно почти сразу после рождения детеныша. Убегая, самка спасает себя – полноценного производителя, оплодотворенную яйцеклетку и иногда висящего на соске новорожденного детеныша.
Однако при этом большую часть ответственности за судьбу детеныша, размером и формой напоминающего фасолину, несет уже не мать, а он сам, и прежде всего его головной мозг. А успешное функционирование системы и одновременно ее глубокие структурные преобразования невозможны. Кроме того, у детенышей сумчатых развивается более мощная по сравнению с детенышами плацентарных мускулатура, необходимая для сосания, что тормозит развитие черепной коробки и, соответственно, мозга.
Можно предположить, что именно плацентарность сыграла роль спускового механизма для неуклонного прогрессивного развития мозга
Плацентарность максимально продлила внутриутробный период и тем самым сняла с мозга детеныша ответственность за контроль над развивающимся организмом. Можно предположить, что она сыграла роль спускового механизма для развития центральной нервной системы – одного из важнейших условий для прогрессивной модернизации мозга в индивидуальном и историческом развитии. Именно поэтому плацентарные – бывшие изгои динозавровой биоты – дали в кайнозое огромное разнообразие форм и стали господствующей группой.
Кроме того, на южных материках Земли млекопитающие жили на сравнительно небольших и однородных в физико-географическом смысле территориях в условиях ослабленной конкуренции. На огромных же пространствах северных материков условия обитания и сообщества наземных позвоночных были значительно многообразнее, а конкуренция – жестче. В этой ситуации для эволюционного успеха было недостаточно одних лишь морфологических, структурных преимуществ: уже в раннем кайнозое в группе млекопитающих наблюдается быстрая эволюция головного мозга. Палеонтологическая летопись сохранила прямые свидетельства увеличения и усложнения мозга плацентарных на протяжении кайнозоя, что по-видимому, привело к значительному усложнению поведения млекопитающих.
Сложное поведение и обучаемость в ходе эволюции начинали развиваться во многих группах животных, но только у плацентарных млекопитающих эта важнейшая эволюционная тенденция достигла своего наиболее яркого и полного выражения. В течение кайнозоя постоянно совершенствовалась способность плацентарных передавать информацию от поколения к поколению негенетическим путем. Закономерным итогом этого неуклонного прогрессивного развития мозга стало появление человека разумного.
Агаджанян А. К. Вопросы ранней радиации млекопитающих // Палеонтологический журнал. 2003. № 1. С. 78—91
Татаринов Л. П. Териодонты СССР// Труды Палеонтологического института РАН. М.: Наука, 1974. Т. 143. 250 с.
Татаринов Л. П. Морфологическая эволюция териодонтов и общие проблемы филогенетики. М.: Наука, 1976. 258 с.
Gambaryan P. P., Kielan-Jaworowska Z. Masticatory musculature of Asian taeniolabidoid multitubrculate mammals // Acta Paleontologica Polonica.1995. V. 40. N 1. P. 45—108
Gambaryan P. P., Kielan-Jaworowska Z. Sprawling versus parasagittal stance in multituberculate mammals // Acta Paleontologica Polonica.1997. V. 42. N 1. P. 13—44
Hu Yaoming, Meng Jin, Wang Yuanqing, Li Chuankui. Large Mesozoic mammals fed on young dinosaurua // Nature. 2005. V. 433. P. 149—152
Jenkins F. A. Jr., Schaff Ch. R. The early Cretaceous mammal Gobiconodon (Mammalia, Triconodontia) from the Cloverly Formation in Montana. // J. Vertebrate Paleontology.1988. V. 8. N 1. P. 1—24
Kielan-Jaworowska Z., Gambaryan P. P. Postcranial anatomy and habitas of Asian multituberculate mammals // Fossils and Strata. 1994. N 36. 92 p.
Novacek M. J., Rougier G. W., Wible J. R., McKenna M. C., Dashzeveg D., Horovitz I. Epipubic bones in eutherian mammals from the Late Cretaceous of Mongolia // Nature. 1997. V. 389. P. 483—486
Hurum J. H., Presley R., Kielan-Jaworoeska Z. The middle ear in multituberculate mammals // Acta Paleontologica Polonica.1996. V. 41. N 3. P. 253—275
Rougier G. W., Wible J. R., Novacek M. J. Middle-ear ossicles of the Multituberculate Kryptobaatar from the Mongolian Late Cretaceous: implications for mammaliamorph relationships and the evolution of auditory apparatus // Novitates American Museum. 1996. N 3187. 43 p.
Исследования поддержаны грантом РФФИ №08-04-00483-а, программой Президиума РАН «Происхождение и эволюция биосферы» и программой ОБН РАН «История биоресурсных видов и сообществ млекопитающих как основа оценки их современного состояния и перспектив сохранения в будущем»