Пирамиды продолговатого мозга содержат аксоны
Medulla oblongata имеет вид луковицы, bulbus cerebri (отсюда термин «бульбарные расстройства»); верхний расширенный конец граничит с мостом, а нижней границей служит место выхода корешков I пары шейных нервов или уровень большого отверстия затылочной кости.
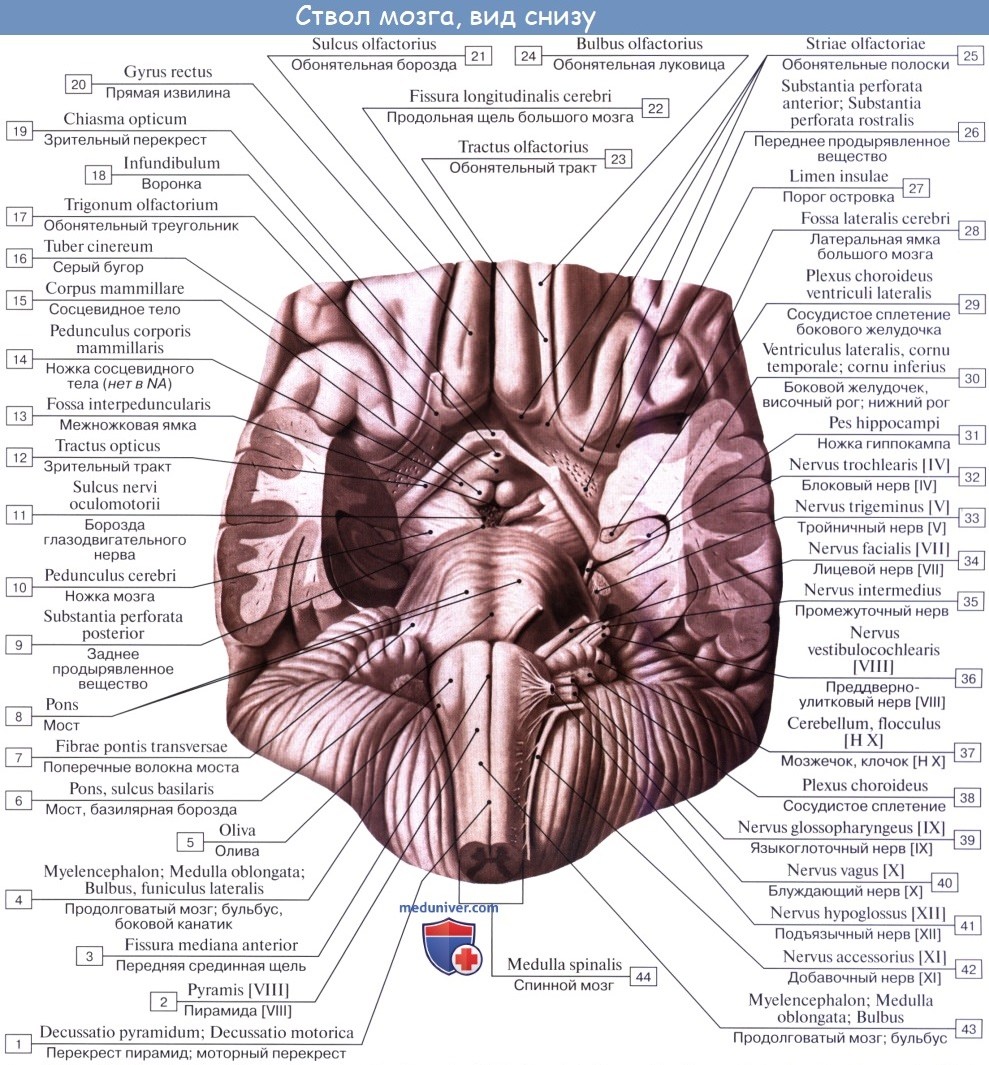
1. На передней (вентральной) поверхности продолговатого мозга по средней линии проходит fissura mediana anterior, составляющая продолжение одноименной борозды спинного мозга. По бокам ее на той и другой стороне находятся два продольных тяжа — пирамиды, pyramides medullae oblongatae, которые как бы продолжаются в передние канатики спинного мозга.
Составляющие пирамиды пучки нервных волокон частью перекрещиваются в глубине fissura mediana anterior с аналогичными волокнами противоположной стороны — decussatio pyramidum, после чего спускаются в боковом канатике на другой стороне спинного мозга — tractus corticospinal (pyramidalis) lateralis, частью остаются неперекрещенными и спускаются в переднем канатике спинного мозга на своей стороне — tractus corticospinalis (pyramidalis) anterior.
Пирамиды отсутствуют у низших позвоночных и появляются по мере развития новой коры; поэтому они наиболее развиты у человека, так как пирамидные волокна соединяют кору большого мозга, достигшую у человека наивысшего развития, с ядрами черепных нервов и передними рогами спинного мозга,
Латерально от пирамиды лежит овальное возвышение — оливa, oliva, которая отделена от пирамиды бороздкой, sulcus anterolateral.
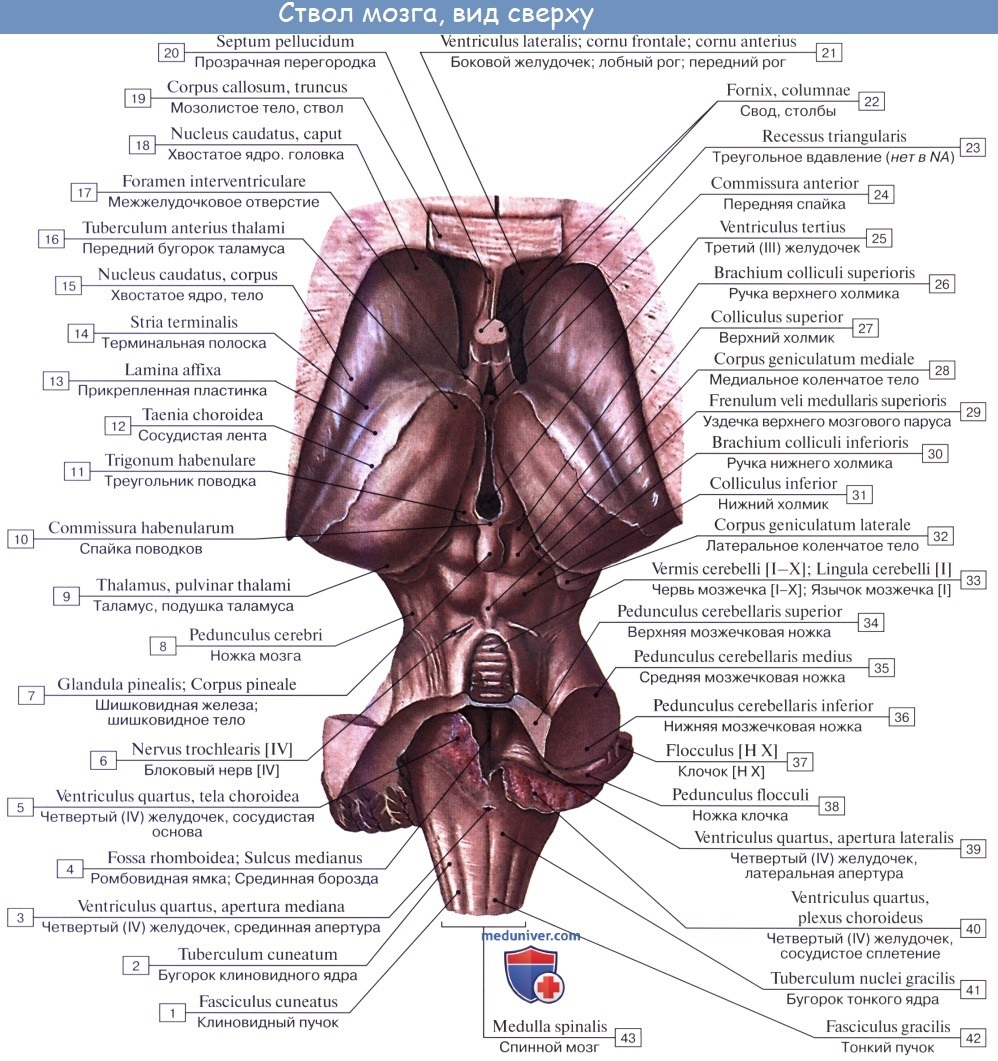
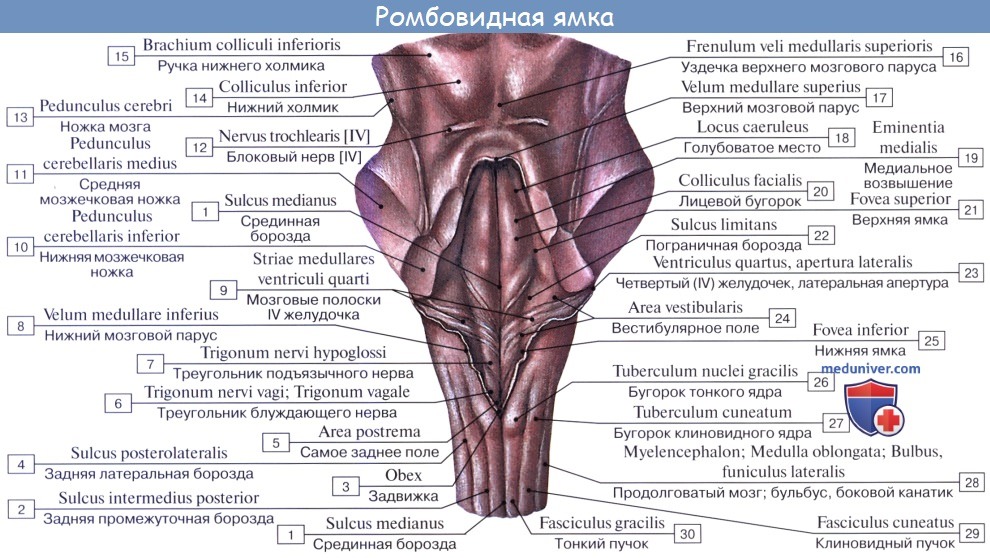
2. На задней (дорсальной) поверхности продолговатого мозга тянется sulcus medianus posterior — непосредственное продолжение одноименной борозды спинного мозга. По бокам ее лежат задние канатики, ограниченные латерально с той и другой стороны слабо выраженной sulcus posterolaterals. По направлению кверху задние канатики расходятся в стороны и идут к мозжечку, входя в состав его нижних ножек, pedunculi cerebellares inferiores, окаймляющих снизу ромбовидную ямку.
Каждый задний канатик подразделяется при помощи промежуточной борозды на медиальный, fasciculus gracilis, и латеральный, fasciculus cuneatus. У нижнего угла ромбовидной ямки тонкий и клиновидный пучки приобретают утолщения — tuberculum gracilum и tuberculum cuneatum. Эти утолщения обусловлены соименными с пучками ядрами серого вещества, nucleus gracilis и nucleus cuneatus.
В названных ядрах оканчиваются проходящие в задних канатиках восходящие волокна спинного мозга (тонкий и клиновидный пучки). Латеральная поверхность продолговатого мозга, находящаяся между sulci posterolateralis et anterolateralis, соответствует боковому канатику. Из sulcus posterolateralis позади оливы выходят XI, X и IX пары черепных нервов. В состав продолговатого мозга входит нижняя часть ромбовидной ямки.
Внутреннее строение продолговатого мозга. Продолговатый мозг возник в связи с развитием органов гравитации и слуха, а также в связи с жаберным аппаратом, имеющим отношение к дыханию и кровообращению. Поэтому в нем заложены ядра серого вещества, имеющие отношение к равновесию, координации движений, а также к регуляции обмена веществ, дыхания и кровообращения.
1. Nucleus olivaris, ядро оливы, имеет вид извитой пластинки серого вещества, открытой медиально (hilus), и обусловливает снаружи выпячивание оливы. Оно связано с зубчатым ядром мозжечка и является промежуточным ядром равновесия, наиболее выраженным у человека, вертикальное положение которого нуждается в совершенном аппарате гравитации. (Встречается еще nucleus olivaris accessorius medialis.)
2. Formatio reticularis, ретикулярная формация, образующаяся из переплетения нервных волокон и лежащих между ними нервных клеток.
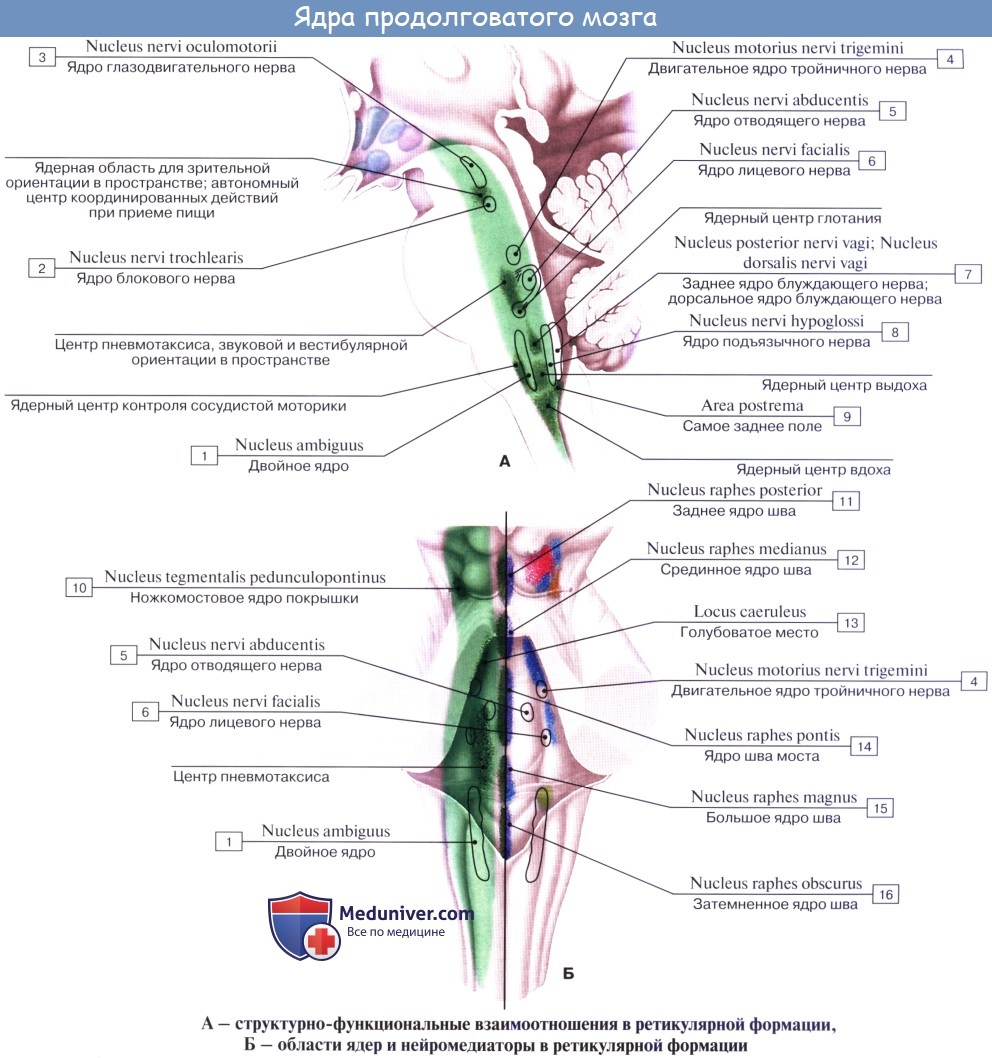
3. Ядра четырех пар нижних черепных нервов (XII —IX), имеющие отношение к иннервации производных жаберного аппарата и внутренностей.
Белое вещество продолговатого мозга содержит длинные и короткие волокна. К длинным относятся проходящие транзитно в передние канатики спинного мозга нисходящие пирамидные пути, частью перекрещивающиеся в области пирамид. Кроме того, в ядрах задних канатиков (nuclei gracilis et cuneatus) находятся тела вторых нейронов восходящих чувствительных путей. Их отростки идут от продолговатого мозга к таламусу, tractus bulbothalamicus.
Волокна этого пучка образуют медиальную петлю, lemniscus medialis, которая в продолговатом мозге совершает перекрест, decussatio lemniscorum, и в виде пучка волокон, расположенных дорсальнее пирамид, между оливами — межоливныи петлевой слой — идет далее. Таким образом, в продолговатом мозге имеется два перекрестка длинных проводящих путей: вентральный двигательный, decussatio pyramidum, и дорсальный чувствительный, decussatio lemniscorum.
К коротким путям относятся пучки нервных волокон, соединяющие между собой отдельные ядра серого вещества, а также ядра продолговатого мозга с соседними отделами головного мозга. Среди них следует отметить tractus olivocerebellaris и лежащий дорсально от межоливного слоя fasciculus longitudinalis medialis.
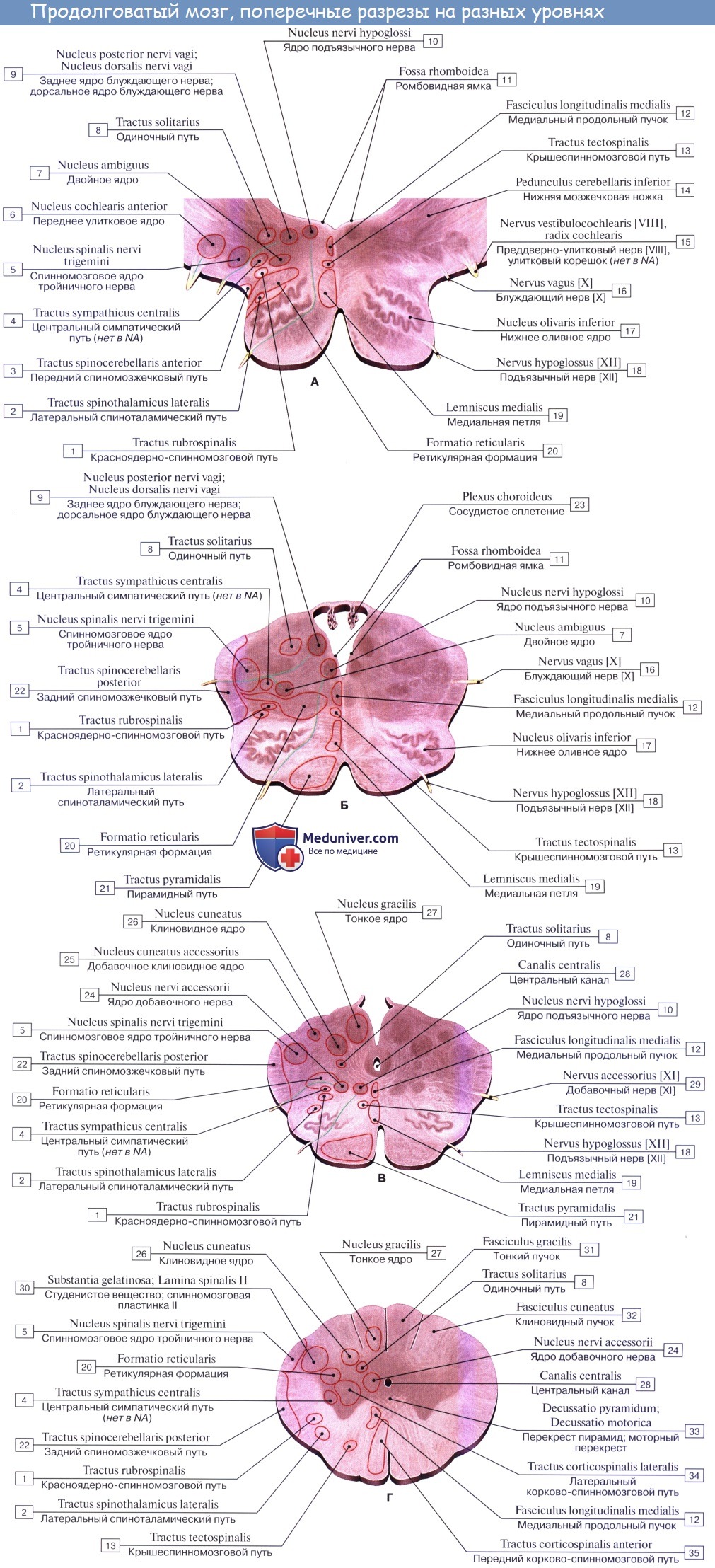
Топографические взаимоотношения главнейших образований продолговатого мозга видны на поперечном срезе, проведенном на уровне олив. Отходящие от ядер подъязычного и блуждающего нервов корешки делят продолговатый мозг на той и другой стороне на три области: заднюю, боковую и переднюю. В задней лежат ядра заднего канатика и нижние ножки мозжечка, в боковой — ядро оливы и formatio reticularis и в передней — пирамиды.
Пирамиды продолговатого мозга содержат аксоны
Существуют следующие нисходящие проводящие пути:
• корково-спинномозговой проводящий путь (пирамидный проводящий путь);
• ретикуло-спинномозговой проводящий путь (экстрапира-мидный путь);
• преддверно-спинномозговой проводящий путь;
• покрышечно-спинномозговой проводящий путь;
• шовно-спинномозговой проводящий путь;
• проводящие пути аминергических систем ЦНС;
• проводящие пути вегетативной нервной системы.
Корково-спинномозговой проводящий путь
Корково-спинномозговой проводящий путь представляет собой крупный проводящий путь произвольной двигательной активности. Около 40 % его волокон начинается из первичной моторной коры прецентральной извилины. Остальные волокна берут начало из дополнительной моторной области на медиальной стороне полушария, премоторной коры головного мозга на латеральной стороне полушария, соматической сенсорной коры, коры теменной доли и коры поясной извилины. Волокна от двух вышеупомянутых сенсорных центров заканчиваются на чувствительных ядрах ствола головного мозга и спинного мозга, где они регулируют передачу чувствительных импульсов.
Корково-спинномозговой проводящий путь спускается вниз через лучистый венец и заднюю ножку внутренней капсулы к стволу головного мозга. Затем он проходит в ножке (головного мозга) на уровне среднего мозга и базилярной части моста, достигая продолговатого мозга. Здесь он образует пирамиду (отсюда название — пирамидный проводящий путь).
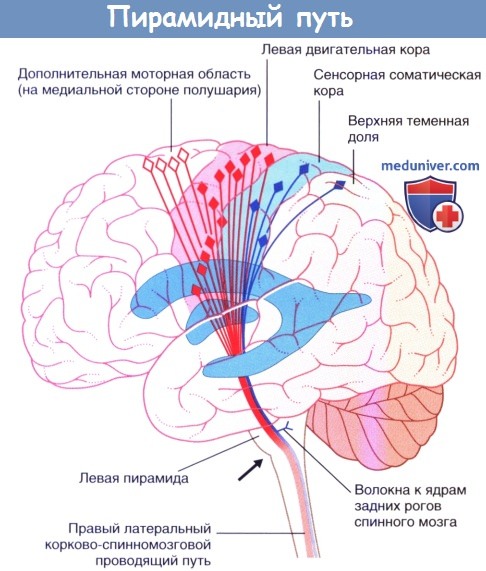 Демонстрация хода волокон пирамидного пути с левой стороны.
Демонстрация хода волокон пирамидного пути с левой стороны.
Дополнительная моторная область на медиальной стороне полушария.
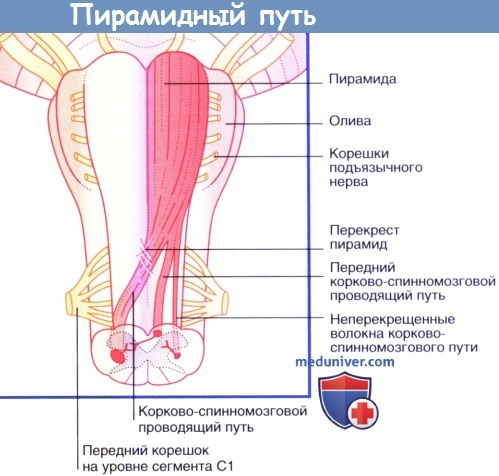
Стрелкой показан уровень перекреста пирамид. Чувствительные нейроны выделены синим цветом. 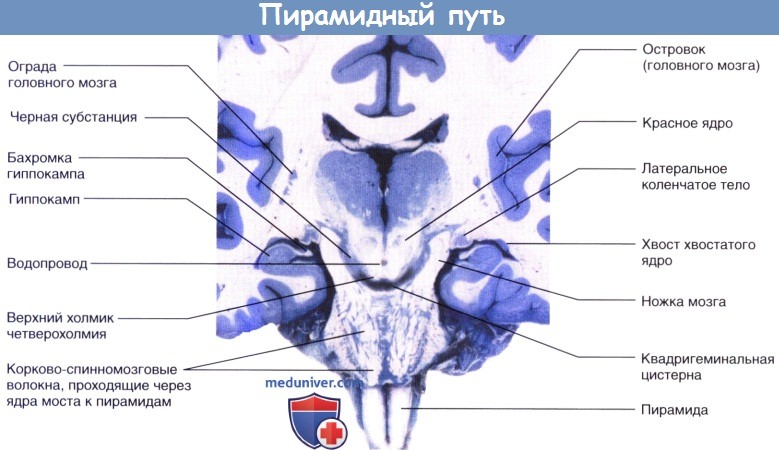 Коронарный срез бальзамированного головного мозга пациента с последующей обработкой сульфатом меди (окраска по Маллигану),
Коронарный срез бальзамированного головного мозга пациента с последующей обработкой сульфатом меди (окраска по Маллигану),
демонстрирующий неокрашенные корково-спинномозговые волокна, идущие через ядра моста в сторону пирамид.
Характеристика волокон корково-спинномозгового пути выше уровня спинномозгового перехода:
• около 80 % (70-90 %) волокон переходят на противоположную сторону на уровне перекреста пирамид;
• эти волокна спускаются по противоположной стороне спинного мозга и составляют латеральный корково-спинномозговой проводящий путь (перекрещивающийся корково-спинномозговой проводящий путь); оставшиеся 20 % волокон не перекрещиваются и продолжают спускаться вниз в передней части спинного мозга;
• половина из этих неперекрещивающихся волокон вступает в передний/вентральный корково-спинномозговой проводящий путь и располагается в вентральном/переднем канатике спинного мозга на шейном и верхнем грудном уровнях; данные волокна переходят на противоположную сторону на уровне белой спайки и иннервируют мышцы передней и задней стенок брюшной полости;
• другая половина вступает в латеральный корково-спинномозговой проводящий путь на своей половине спинного мозга.
Считают, что корково-спинномозговой проводящий путь содержит около 1 млн. нервных волокон. Средняя скорость проведения импульса составляет 60 м/с, что указывает на средний диаметр волокна, равный 10 мкм («правило шести»). Около 3 % волокон — очень крупные (до 20 мкм); они отходят от гигантских нейронов (клетки Беца), расположенных в основном в области двигательной коры, отвечающей за иннервацию нижних конечностей. Все волокна корково-спинномозгового пути — возбуждающие и в качестве медиатора используют глутамат.
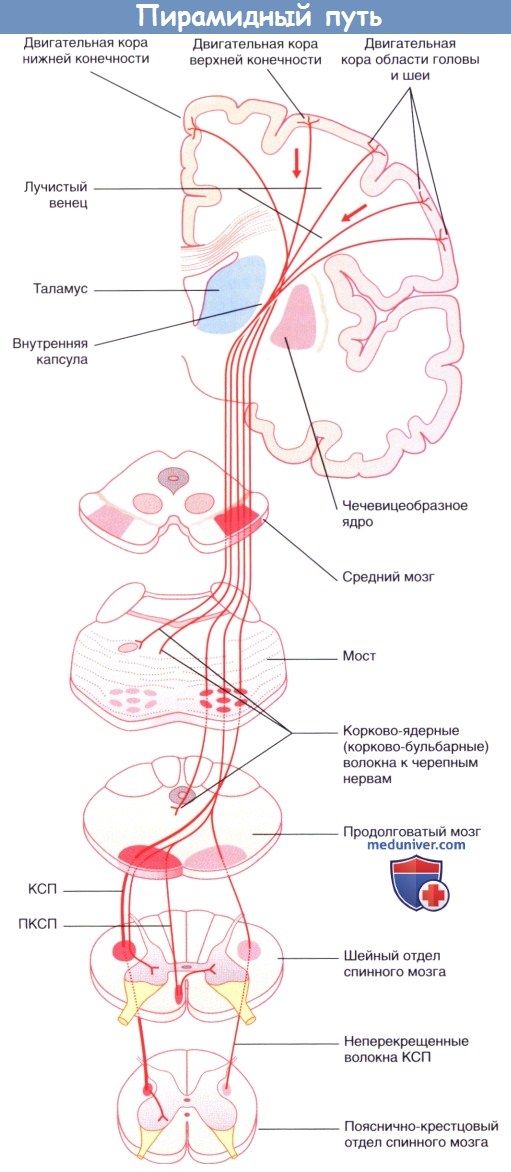 Пирамидный проводящий путь.
Пирамидный проводящий путь.
КСП — корково-спинномозговой проводящий путь;
ПКСТ — передний корково-спинномозговой проводящий путь;
ЛКСП — латеральный корково-спинномозговой проводящий путь.
Обратите внимание: показан только двигательный компонент; компоненты теменной доли опущены.
Клетки-мишени латерального корково-спинномозгового пути:
а) Мотонейроны дистальных отделов конечностей. В передних рогах серого вещества спинного мозга аксоны латерального корково-спинномозгового пути могут непосредственно образовывать синапсы на дендритах α- и γ-мотонейронов, иннервирующих мышцы конечностей, особенно верхних (однако, как правило, это происходит через интернейроны в пределах серого вещества спинного мозга). Отдельные аксоны латерального корково-спинномозгового пути могут активировать «большие» или «малые» двигательные единицы.
Двигательная единица — это комплекс, состоящий из нейрона переднего рога спинного мозга и всех мышечных волокон, которые этот нейрон иннервирует. Нейроны малых двигательных единиц избирательно иннервируют небольшое количество мышечных волокон и участвуют в выполнении тонких и точных движений (например, при игре на пианино). Нейроны переднего рога, иннервирующие крупные мышцы (например, большую ягодичную мышцу), способны по отдельности вызвать сокращение сотни мышечных клеток сразу, так эти мышцы отвечают за грубые и простые движения.
Уникальное свойство этих корковомотонейронных волокон латерального корково-спинномозгового пути демонстрирует понятие «фракционирования», относящееся к переменной активности интернейронов, в результате чего небольшие группы нейронов могут быть избирательно активированы для выполнения конкретной общей функции. Это легко показать на указательном пальце, который может быть согнут или разогнут независимо от положения других пальцев (хотя три из его длинных сухожилий имеют общее начало с мышечным ложем всех четырех пальцев).
Фракционирование имеет большое значение при выполнении привычных движений, таких как застегивание пальто или завязывание шнурков. Травматическое или другое повреждение корковомотонейронной системы на любом уровне влечет за собой утрату навыков выполнения привычных движений, которые затем редко поддаются восстановлению.
При выполнении данных движений α- и γ-мотонейроны активируются совместно через латеральный корково-спинномозговой проводящий путь таким образом, что веретена мышц, первично задействованных в движении, посылают импульсы об активном растяжении, а веретена мышц-антагонистов — о пассивном растяжении.
 Продолговатый мозг и верхние отделы спинного мозга, вид спереди.
Продолговатый мозг и верхние отделы спинного мозга, вид спереди.
Продемонстрированы три группы нервных волокон левой пирамиды.
б) Клетки Реншоу. Функции синапсов латерального корково-спинномозгового пути на клетках Реншоу довольно многочисленны, так как торможение на некоторых клеточных синапсах главным образом происходит за счет интернейронов типа Iа; на других синапсах данную функцию выполняют клетки Реншоу. Вероятно, наиболее важная функция — контроль совместного сокращения основных движущих мышц и их антагонистов для фиксации одного или нескольких суставов, например при работе с кухонным ножом или лопатой. Совместное сокращение происходит за счет инактивации ингибирующих интернейронов Iа клетками Реншоу.
в) Возбуждающие интернейроны. Латеральный корково-спинно-мозговой проводящий путь влияет на деятельность двигательных нейронов, расположенных в средней части серого вещества и в основании переднего рога спинного мозга, иннервирующих осевые (позвоночные) мышцы и мышцы проксимальных отделов конечностей посредством возбуждающих интернейронов. г) la-ингибирующие интернейроны. Эти нейроны также расположены в средней части серого вещества спинного мозга и активируются латеральным корково-спинномозговым путем в первую очередь при совершении произвольных движений.
Активность Ia-интернейронов способствует расслаблению мышц-антагонистов до того, как начнут сокращаться мышцы-агонисты. Кроме того, они вызывают рефрактерность мотонейронов мышц-антагонистов к стимуляции афферентами нервно-мышечного веретена при их пассивном растяжении во время движения. Последовательность процессов при произвольном сгибания коленного сустава показана на рисунке ниже.
(Обратите внимание на терминологию: в спокойном положении стоя колени человека «закрыты» в небольшом переразгибании, а четырехглавая мышца бедра находится в неактивном состоянии, о чем свидетельствует «свободное» положение надколенника. При попытке сгибания одного или обоих колен происходит подергивание четырехглавой мышцы бедра в ответ на пассивное растяжение в ней десятков мышечных веретен. Поскольку таким образом происходит сопротивление сгибанию, рефлекс называют рефлексом сопротивления.
С другой стороны, во время произвольного сгибания коленного сустава мышцы способствуют данному движению с помощью такого же механизма, но уже через рефлекс помощи. Изменение знака с отрицательного на положительный называют рефлексом перемены направления.)
д) Пресинаптические ингибиторные нейроны, обеспечивающие рефлекс растяжения. Рассмотрим движения спринтера. На каждом шаге сила тяжести тянет его тело вниз, на выпрямленное четырехглавой мышцей колено. В момент соприкосновения с землей все нервно-мышечные веретена в сокращенной четырехглавой мышце резко растягиваются, в результате чего возникает опасность разрыва мышцы. Сухожильный орган Гольджи обеспечивает некоторую защиту посредством внутреннего торможения, однако основной защитный механизм обеспечивает латеральный корково-спинномозговой путь через пресинаптическое торможение афферентов веретен вблизи их контакта с мотонейронами.
В то же время удлинение паузы до ахиллового рефлекса служит преимуществом в этой ситуации, так как происходит восстановление мотонейронов, иннервирующих заднюю часть голени, для следующего рывка. Предполагают, что степень подавления рефлекса растяжения со стороны латерального корково-спинномозгового пути зависит от конкретных движений.
е) Пресинаптическое ингибирование чувствительных нейронов первого порядка. В заднем роге серого вещества спинного мозга существует некоторое подавление передачи чувствительных импульсов в спиноталамический проводящий путь при совершении произвольных движений. Это происходит путем активации синапсов, образованных ингибирующими вставочными нейронами и первичными чувствительными нервными окончаниями.
Еще более тонкую регуляцию наблюдают на уровне тонкого и клиновидного ядер, где волокна пирамидного пути (после пересечения) способны усиливать передачу чувствительных импульсов во время медленных аккуратных движений или ослаблять ее во время совершения быстрых движений.
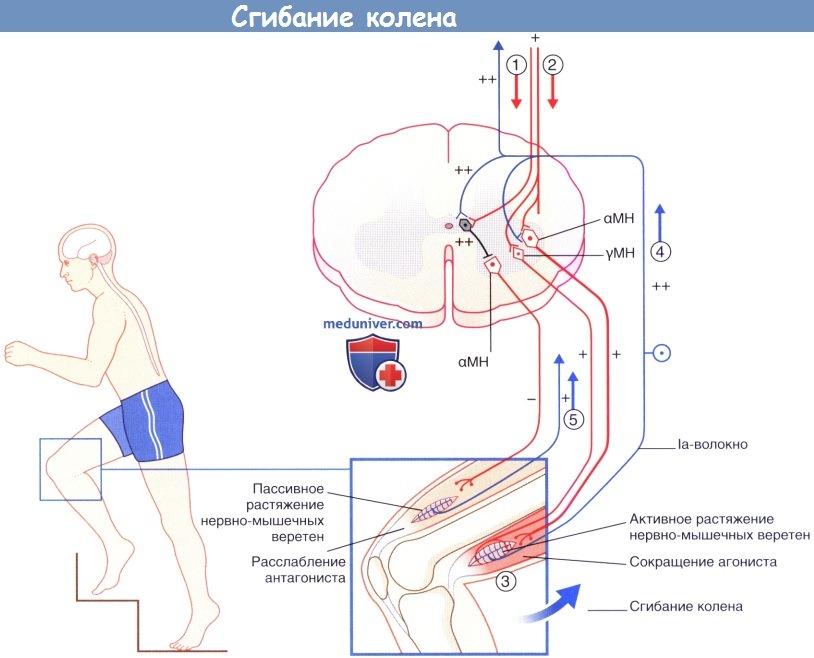 Последовательность событий при выполнении произвольного движения (сгибания колена). МН — мотонейроны.
Последовательность событий при выполнении произвольного движения (сгибания колена). МН — мотонейроны.
(1) Активация la интернейронов ингибирует их антагонисты-α-мотонейроны.
(2) Активация агонистов α- и γ-мотонейронов.
(3) Активация экстрафузальных и интрафузальных мышечных волокон.
(4) Импульсация от активно растянутых нервно-мышечных веретен увеличивает активность агониста а-мотонейрона и снижает активность его антагонистов.
(5) Iа-волокна от пассивно растянутых нервно-мышечных веретен-антагонистов направляются к соответствующим рефрактерным а-мотонейронам.
Обратите внимание: последовательность «γ-мотонейронон—Ia-волокно—α-мотонейрон» образует γ-петлю.
Редактор: Искандер Милевски. Дата публикации: 15.11.2018
Продолговатый мозг
Продолговатый мозг, medulla oblongata (myelencephalon), является непосредственным продолжением спинного мозга кверху. Продолговатый мозг имеет форму конуса или луковицы, bulbus (отсюда термин «бульбарные расстройства»). Он сочетает в себе особенности строения спинного и головного мозга, поэтому и называется myelencephalon.
Границы. Продолговатый мозг внизу обращен к спинному мозгу, вверху – к мосту и мозжечку. Верхняя граница продолговатого мозга на вентральной поверхности головного мозга проходит по нижнему краю моста (бульбарно-мостовая борозда), на дорсальной поверхности соответствует мозговым полоскам IV желудочка, которые делят дно его на верхнюю и нижнюю части.
Граница между продолговатым и спинным мозгом соответствует нижнему уровню перекреста пирамид (уровень большого затылочного отверстия) или месту выхода из мозга первого шейного спинномозгового нерва.
Внешнее строение. Продольный размер продолговатого мозга составляет 2,5 – 3,2 см, поперечный – в среднем 1,5 см, переднезадний – до 1 см. Мал золотник, да дорог.
В продолговатом мозге выделяют вентральную, дорсальную и две боковые поверхности, которые разделены бороздами. Борозды продолговатого мозга являются непосредственным продолжением борозд спинного мозга и носят те же названия. Передняя срединная щель, fissura mediana anterior, задняя срединная борозда, sulcus medianus dorsalis, переднелатеральная борозда, sulcus ventrolateralis и заднелатеральная борозда, sulcus dorsolateralis.
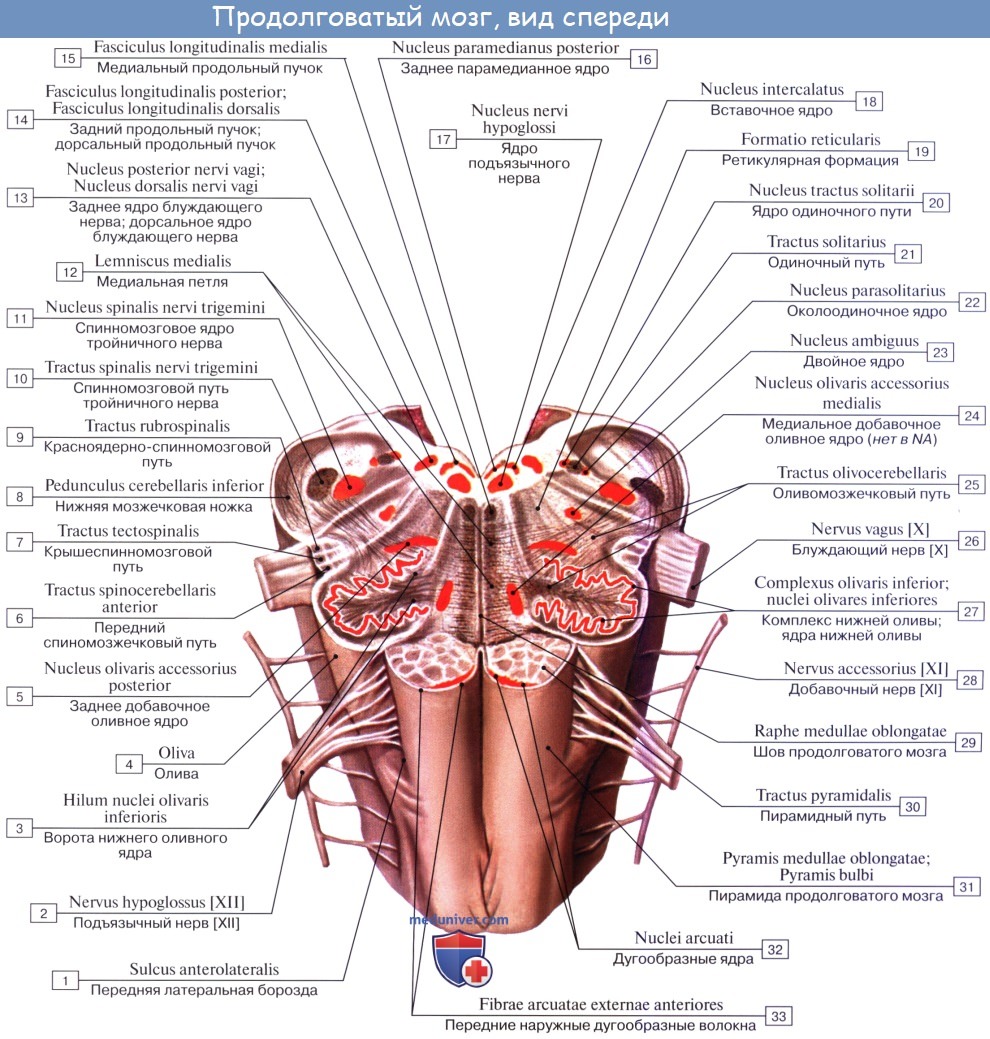
Вентральная поверхность продолговатого мозга расположена на скате и занимает его нижний отдел до большого затылочного отверстия. На передней поверхности продолговатого мозга по обе стороны от передней срединной щели расположены конусообразные валики, постепенно суживающиеся книзу – пирамиды, pyramides. Они являются непосредственным продолжением передних канатиков спинного мозга. В нижней части продолговатого мозга часть волокон, составляющих пирамиды, переходят на противоположную сторону и вступают в боковые канатики спинного мозга (латеральный корково-спинномозговой путь). Этот переход волокон получил название перекреста пирамид, decussatio pyramidum. Место перекреста также служит анатомической границей между продолговатым и спинным мозгом. Остальная часть волокон не переходит на противоположную сторону, т.е. не перекрещивается, и спускается вниз в составе передних канатиков спинного мозга своей стороны (передний корково-спинномозговой путь).
Сбоку от каждой пирамиды продолговатого мозга находится овальное возвышение – олива, oliva. Они являются непосредственным продолжением боковых канатиков спинного мозга. Олива отделена от пирамиды при помощи переднелатеральной борозды, из которой выходят корешки (6 – 10) подъязычного нерва (XII пара).
Дорсальнее каждой оливы, из заднелатеральной борозды, которая здесь называется позадиоливной бороздой, sulcus retroolivaris, выходят корешки языкоглоточного, блуждающего и добавочного нервов (IX, X и XI пары).
На дорсальной поверхности продолговатого мозга, по бокам от задней срединной борозды, проходят задние канатики, ограниченные латерально заднелатеральной бороздой. По направлению кверху задние канатики расходятся в стороны и идут к мозжечку, входя в состав его нижних ножек, окаймляющих снизу ромбовидную ямку. Каждый канатик при помощи промежуточной борозды разделяется на два пучка: тонкий и клиновидный. У нижнего угла ромбовидной ямки тонкий и клиновидный пучки заканчиваются утолщениями. Лежащий более медиально тонкий пучок, fasciculus gracilis, расширяясь, образует бугорок тонкого ядра, tuberculum gracile. Латеральнее располагается клиновидный пучок, fasciculus cuneatus, который сбоку от бугорка тонкого пучка образует бугорок клиновидного ядра, tuberculum cuneatum. Внутри этих бугорков находятся одноименные ядра, nucleus gracilis et cuneatus. В этих ядрах тонкий и клиновидный пучки спинного мозга заканчиваются.
Внутреннее строение. На поперечных разрезах продолговатого мозга на уровне пирамид можно определить, что каждая пирамида представляет собой комплекс пучков, которые частично взаимно перекрещиваются. Перекрещивающиеся волокна переходят в боковой канатик спинного мозга с противоположной стороны и далее следуют как латеральный корково-спинномозговой путь. Меньшая часть пучков, не входящая в перекрест, следует в системе переднего канатика спинного мозга как передний корково-спинномозговой путь. Оба эти пути объединяют под названием пирамидный путь. Таким образом, вентральные отделы продолговатого мозга представлены нисходящими двигательными пирамидными путями.
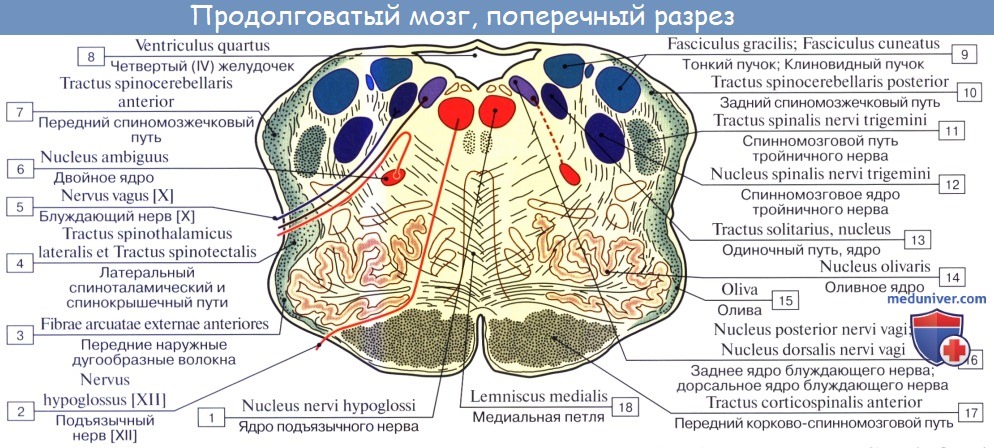
На поперечном разрезе продолговатого мозга, на уровне олив, видны скопления белого и серого вещества. Скопление белого вещества, которое окружает серое вещество, называется оливным плащом, amiculum olivare. Наибольшее из скоплений серого вещества имеет подковообразную форму и представлено нижними оливными ядрами и добавочными оливными ядрами (задним и медиальным).
Нижние оливные ядра, nuclei olivares inferiores изогнуты таким образом, что их ворота, hilum nuclei olivares caudales, открыты медиально и вверх. Из этих ворот выходят волокна оливомозжечкового пути, tractus olivocerebellaris. Ядра оливы связаны с зубчатым ядром мозжечка и участвуют в регуляции равновесия тела.
Между нижними оливными ядрами располагается так называемый межоливный слой, представленный внутренними дугообразными волокнами – отростками клеток, лежащих в тонком и клиновидном ядрах. Эти волокна в совокупности формируют медиальную петлю, lemniscus medialis. Т.е. медиальная петля является как бы продолжением пучков Голля и Бурдаха в пределах головного мозга. Волокна медиальной петли принадлежат проприоцептивному пути коркового направления и образуют в продолговатом мозге перекрест медиальных петель, decussatio lemniscorum medialium. Таким образом, в продолговатом мозге имеется 2 перекреста проводящих путей: вентральный двигательный, decussatio pyramidum, и дорсальный чувствительный, decussatio lemniscorum.
В продолговатом мозге также залегают ядра IX, X, XI и XII пар черепных нервов.
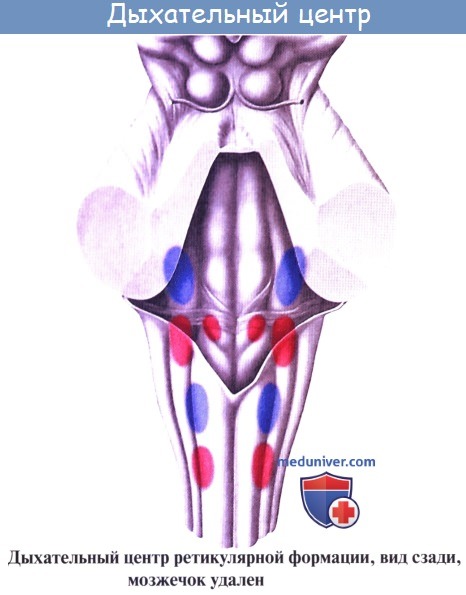
В продолговатом мозге находятся такие жизненно важные центры как дыхательный и сосудодвигательный, а также центр полового возбуждения.
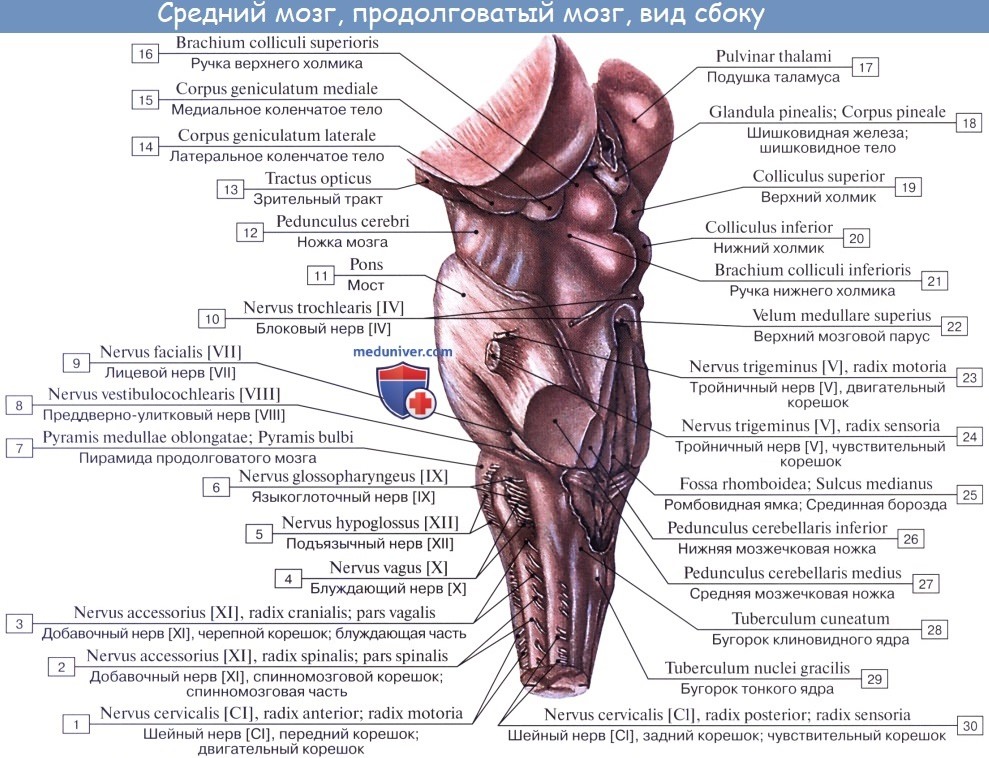
Если посмотреть сбоку, то мы увидим, что корешки подъязычного нерва спереди, блуждающего, языкоглоточного и добавочного нервов сзади делят продолговатый мозг на 3 условные области с каждой стороны: переднюю, боковую и заднюю. В задней области лежат тонкое и клиновидное ядро, в боковой – ядра оливы и ретикулярная формация и в передней – пирамиды.
Задний мозг, metencephalon, включает мост, расположенный спереди (вентрально) и мозжечок, который находится позади моста.
Мост, pons, (варолиев мост) имеет вид поперечного валика, расположенного непосредственно над продолговатым мозгом. На вентральной поверхности ствола мозга вверху он граничит со средним мозгом (с его ножками), а внизу – с продолговатым мозгом, от которого отделяется посредством бульбарно-мостовой борозды, sulcus bulbopontinus. Латеральной границей моста служит условная линия, проведенная через корешки тройничного и лицевого нервов, тройнично-лицевая линия, linea trigeminofacialis. Латеральнее этой линии мост переходит в средние мозжечковые ножки.
Внешнее строение. Дорсальная поверхность моста не видна снаружи, т.к. накрыта мозжечком. Ее можно увидеть, если удалить мозжечок. Она обращена в сторону IV желудочка и участвует в образовании ромбовидной ямки, той ее части, которая располагается кверху от мозговых полосок четвертого желудочка.
Вентральная поверхность моста, которая в полости черепа прилежит к верхнему отделу ската, clivus, имеет волокнистое строение, причем волокна идут поперечно и в латеральном направлении с каждой стороны переходят в среднюю мозжечковую ножку, pedunculus cerbellaris medius, уходящую в полушарие мозжечка. В бульбарно-мостовой борозде, отделяющей мост от пирамид продолговатого мозга, выходят корешки правого и левого отводящих нервов (VI пара). В латеральной части этой борозды видны корешки лицевого (VII пара) и преддверно-улиткового нервов (VIII пара).
По средней линии вентральной поверхности проходит базилярная борозда, sulcus basilaris, в которой расположена базилярная артерия.
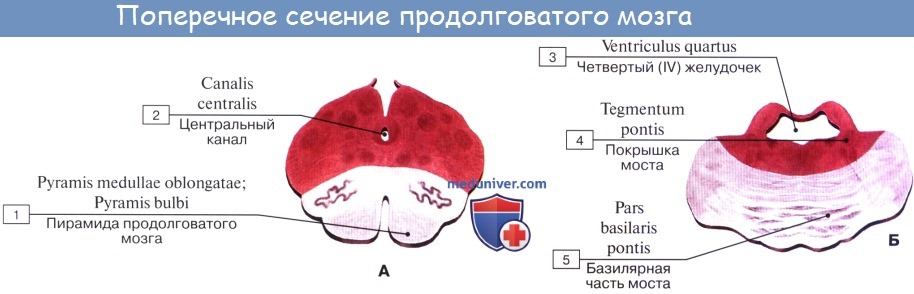
Внутреннее строение. На фронтальных разрезах моста видно, что он состоит из большей вентральной части, pars ventralis pontis, и меньшей дорсальной части или покрышки моста, pars dorsalis (tegmentum pontis). Границей между этими частями является толстый слой поперечных волокон – трапециевидное тело, corpus trapezoideum, волокна которого относятся к слуховому пути.
Между волокнами трапециевидного тела расположены переднее и заднее ядра трапециевидного тела, nuclei corporis trapezoidei ventralis et dorsalis.
В вентральной части моста видны продольные и поперечные нервные волокна. Продольные волокна моста, fibrae pontis longitudinales, принадлежат корково-спинальному и корково-нуклеарному путям. Здесь же имеются и корково-мостовые волокна, fibrae corticopontinae, которые заканчиваются на собственных ядрах моста, nuclei pontis proprii. Отростки нервных клеток собственных ядер моста в свою очередь образуют поперечные волокна моста, fibrae pontis transversae. Эти волокна перекрещиваются с одноименными волокнами с противоположной стороны и образуют средние мозжечковые ножки, pedunculi cerebellares medii. Эти ножки направляются к коре мозжечка.
В дорсальной части (покрышке) моста располагаются ядра V, VI, VII, VIII пар черепных нервов, поверх которых находится выстланное эпендимой дно четвертого желудочка.