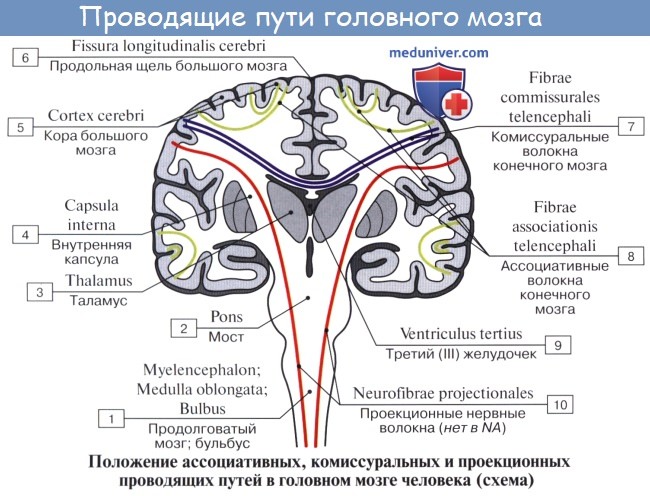
Проекционные волокна связывают мозговую кору частью с thalamus и corpora geniculata, частью с нижележащими отделами центральной нервной системы до спинного мозга включительно. Одни из этих волокон проводят возбуждения центростремительно, по направлению к коре, а другие, наоборот,— центробежно.
Проекционные волокна в белом веществе полушария ближе к коре образуют так называемый лучистый венец, corona radiata, и затем главная часть их сходится во внутреннюю капсулу, о которой упоминалось выше.
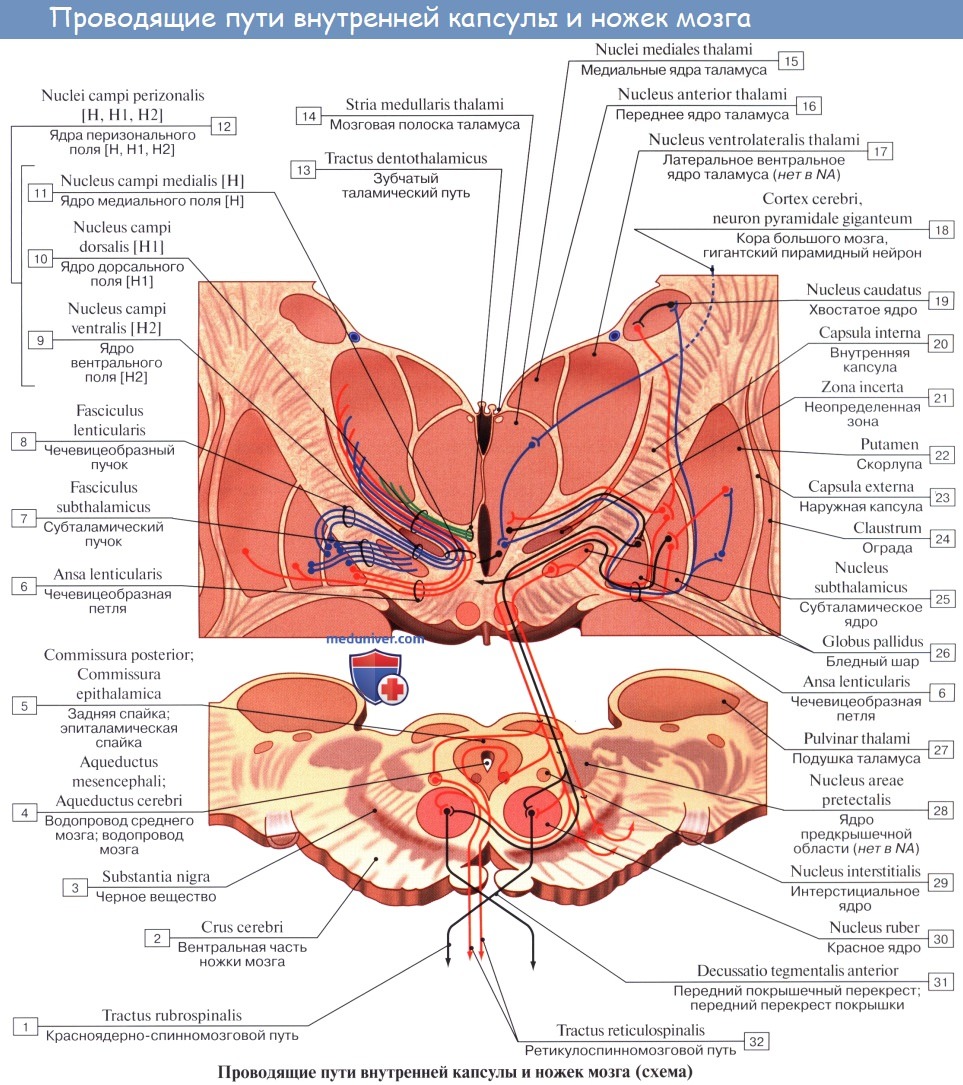
Внутренняя капсула, capsula interna
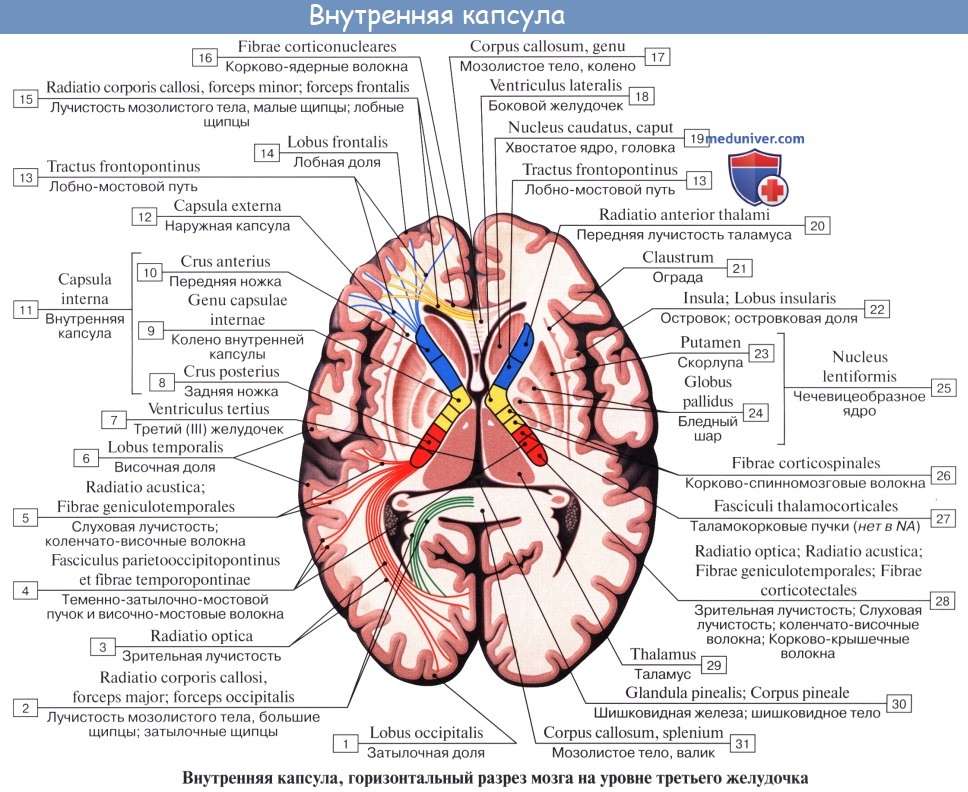
Внутренняя капсула, capsula interna, как было указано, представляет слой белого вещества между nucleus lentiformis, с одной стороны, и хвостатым ядром и таламусом — с другой.
На фронтальном разрезе мозга внутренняя капсула имеет вид косо идущей белой полосы, продолжающейся в ножку мозга. На горизонтальном разрезе она представляется в форме угла, открытого в латеральную сторону; вследствие этого в capsula interna различают переднюю ножку, crus anterius capsulae internae, — между хвостатым ядром и передней половиной внутренней поверхности nucleus lentiformis, заднюю ножку, crus posterius,— между таламусом и задней половиной чечевицеобразного ядра и колено, genu capsulae internae, лежащее на месте перегиба между обеими частями внутренней капсулы. Проекционные волокна по их длине могут быть разделены на следующие системы, начиная с самых длинных:
Видео урок для зубрешки анатомия базальных ядер, внутренней капсулы, белого вещества и волокон полушарий мозга
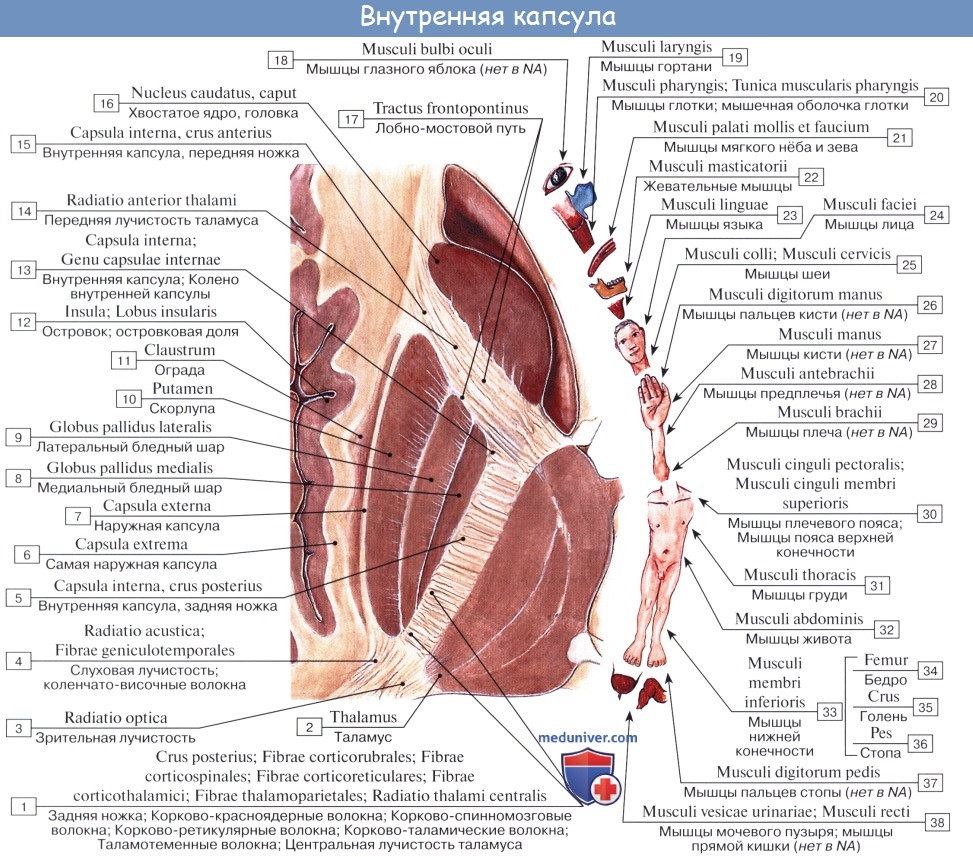
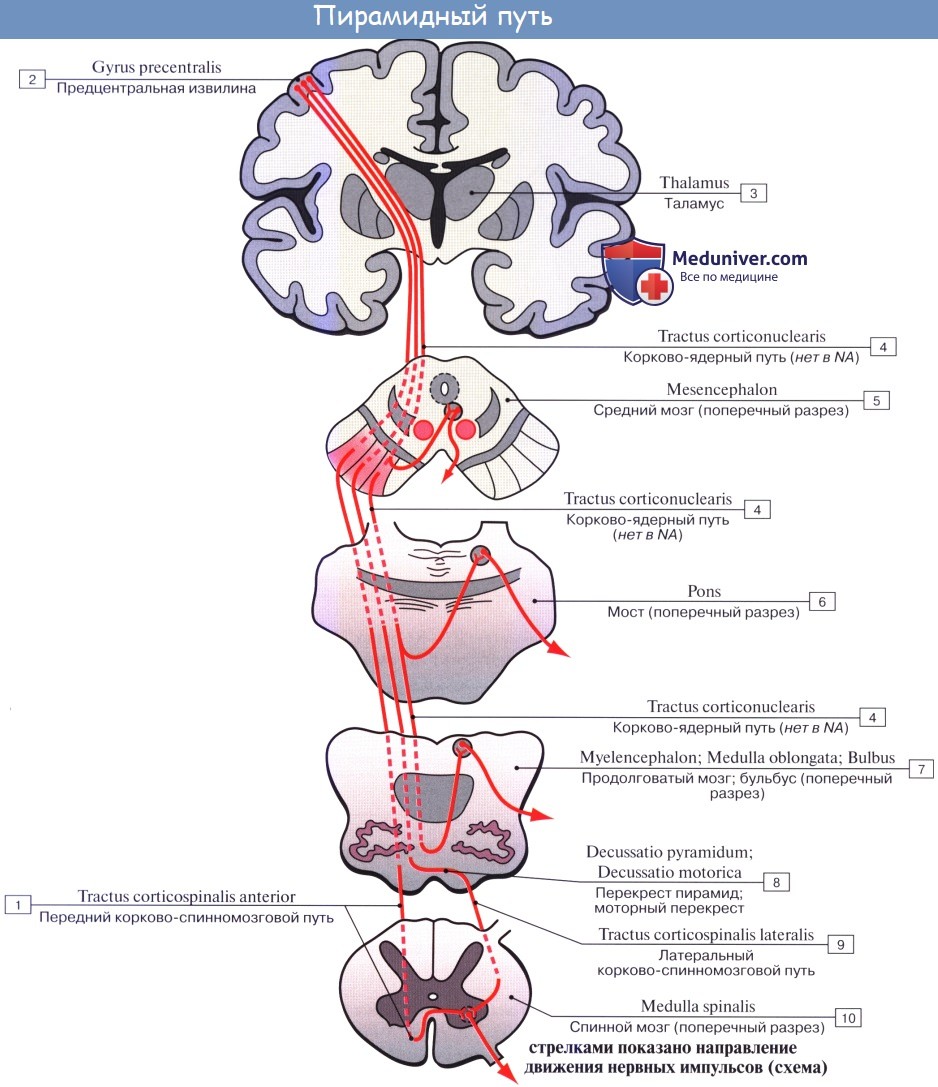
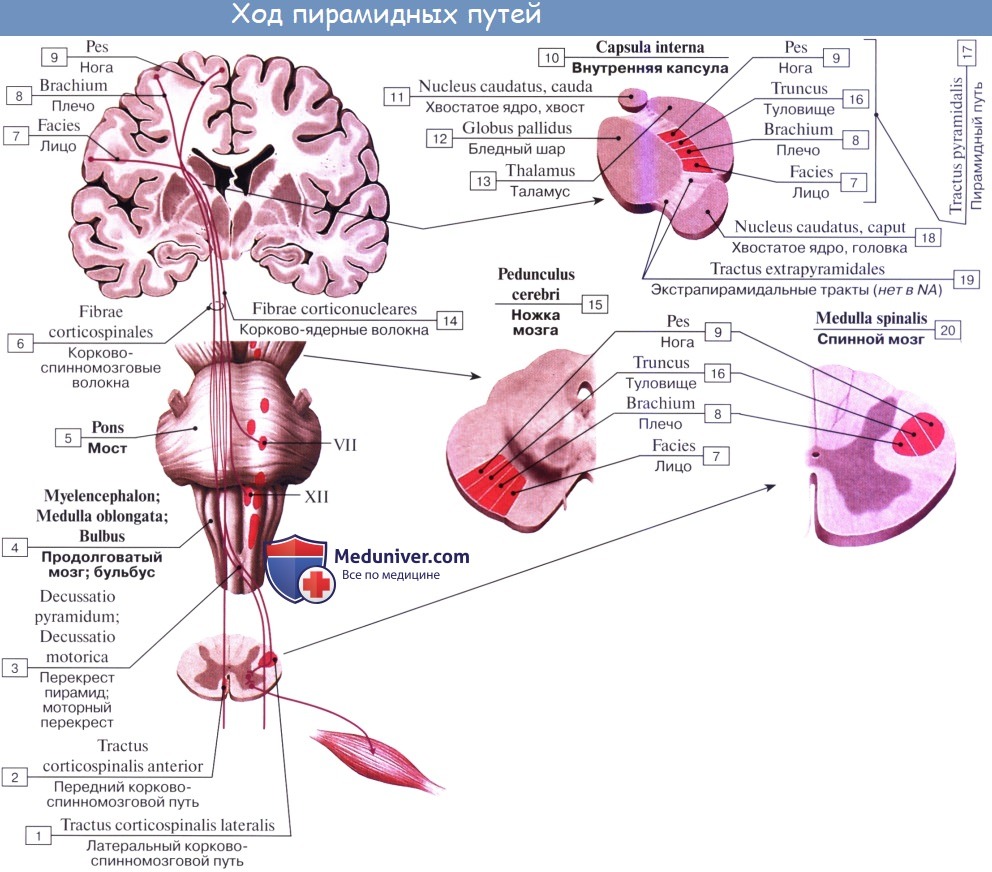
1. Пирамидный путь, tractus corticospinalis (pyramidalis) проводит двигательные волевые импульсы к мышцам туловища и конечностей. Начавшись от пирамидных клеток коры средней и верхней частей предцентральной извилины и lobulus paracentralis, волокна пирамидного пути идут в составе лучистого венца, а затем проходят через внутреннюю капсулу, занимая передние две трети ее задней ножки, причем волокна для верхней конечности идут спереди волокон для нижней конечности. Далее они проходят через ножку мозга, pedunculus cerebri, а оттуда через мост в продолговатый мозг.
Редактор: Искандер Милевски. Дата последнего обновления публикации: 13.8.2020
Проекционные волокна головного мозга
В этой статье на сайте последовательно описаны следующие структуры: таламус, хвостатое и чечевицеобразное ядра, внутренняя капсула; гиппокамп и свод мозга; ассоциативные и комиссуральные волокна; III и боковые желудочки.
а) Таламус, хвостатое и чечевицеобразное ядра, внутренняя капсула. Между дорсальным и вентральным таламусом располагается III желудочек щелевидной формы. В большинстве случаев таламусы соприкасаются, образуя межталамическое сращение. На рисунке ниже показано последовательное расположение таламуса и соседних структур в срединно-боковой проекции. Головка и тело хвостатого ядра соприкасаются с верхней частью таламуса. Хвост хвостатого ядра проходит спереди и снизу от таламуса, не соприкасаясь с ним.
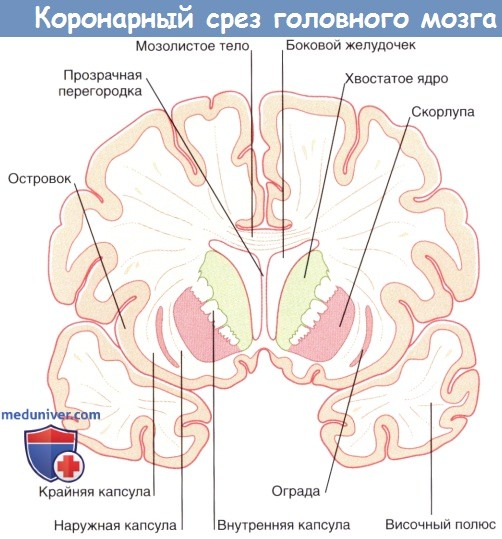
Двояковыпуклое чечевицеобразное ядро состоит из скорлупы и бледного шара. Скорлупа и хвостатое ядро схожи по структуре, и их концы сращены. Эти два образования соединены в задней части пучками серого вещества, располагающимися поперечно по отношению к внутренней капсуле. В связи с этим скорлупа и хвостатое ядро объединены под названием «полосатое тело».
Хвостатое и чечевицеобразное ядра входят в состав базальных ядер. Изначально термин «базальные ядра» применяли к шести скоплениям серого вещества в основании полушарий мозга. В настоящее время это понятие включает четыре ядра, участвующих в обеспечении двигательной иннервации: хвостатое и чечевицеобразное ядра, субталамические ядра промежуточного мозга и черную субстанцию среднего мозга.
На горизонтальном срезе внутренняя капсула имеет зигзагообразную форму. На горизонтальном срезе можно различить пять компонентов внутренней капсулы. 1. Передняя ножка внутренней капсулы, располагающаяся между чечевицеобразным ядром и головкой хвостатого ядра. 2. Колено внутренней капсулы. 3. Задняя ножка внутренней капсулы, располагающаяся между чечевицеобразным ядром и таламусом. 4. Позадичечевичная часть, располагающаяся сзади чечевицеобразного ядра и латерально по отношению к таламусу. 5. Подчечевичная часть (слуховая лучистость).
Через заднюю ножку внутренней капсулы проходит корково-спинномозговой проводящий путь, который также называют пирамидным трактом. Тракт представляет собой пучок волокон, выполняющих одну и ту же функцию. Корково-спинномозговой путь берет начало преимущественно из прецентральной извилины коры. Он спускается и, проходя через лучистый венец, внутреннюю капсулу и ножку среднего мозга (ножку мозга), идет к нижней части ствола мозга, а затем осуществляет перекрест и переходит на противоположную сторону спинного мозга.
Корково-спинномозговой путь — наиболее значимый проводящий путь ЦНС с клинической точки зрения по двум причинам. Во-первых, он обеспечивает все произвольные движения, и повреждения этого пути приводят к ослаблению (парезу) или параличу двигательной мускулатуры. Во-вторых, он удлиняет вертикальную протяженность ЦНС, делая ее уязвимой для заболеваний или травм полушарий и ствола мозга с одной стороны либо для заболеваний или травм спинного мозга с противоположной стороны.
На рисунке ниже изображен коронарный срез передней ножки внутренней капсулы; на рисунке ниже представлена соответствующая МР-томограмма. На рисунке ниже изображен коронарный срез задней ножки внутренней капсулы препарата мозга; на рисунке ниже представлен соответствующая МР-томограмма.
Латеральнее чечевицеобразного ядра расположены наружная капсула, ограда и крайняя капсула.
б) Гиппокамп и свод мозга. Во время эмбрионального развития гиппокамп, играющий важнейшую роль в формировании памяти, сначала можно увидеть сверху от мозолистого тела. У низших млекопитающих основная часть гиппокампа сохраняет свое расположение, тогда как у приматов он перемещается в височную долю по мере ее развития, оставляя за собой след, представленный нервными тяжами белого вещества,— свод мозга. Зрелый гиппокамп формирует дно нижнего (височного) рога бокового желудочка. Зрелый свод мозга состоит из тела свода, расположенного под мозолистым телом, ножек свода, начинающихся от гиппокампа, и двух колонн свода, идущих от свода к промежуточному мозгу. С ножкой и телом свода тесно связана сосудистая щель, через которую сосудистое сплетение заходит в боковой желудочек.
в) Ассоциативные и комиссуральные нервные волокна. Нервные волокна, идущие от коры головного мозга, разделяют на три группы. 1. Ассоциативные волокна, обеспечивающие связи внутри одного полушария. 2. Комиссуральные волокна, соединяющие соответствующие части двух полушарий. 3. Проекционные волокна, идущие к подкорковым ядрам полушарий, ствола мозга и спинного мозга.
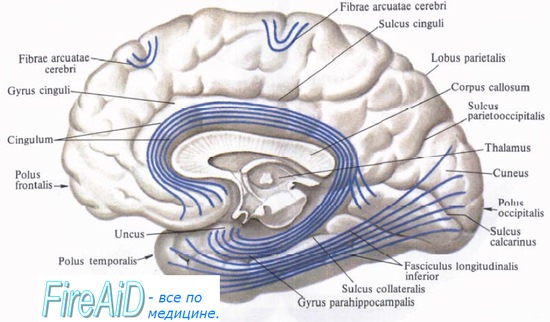
г) Ассоциативные нервные волокна. Извилины в пределах доли соединяются короткими ассоциативными волокнами.
Связи двух долей обеспечивают длинные ассоциативные волокна. К пучкам длинных ассоциативных волокон относят: • верхний продольный пучок, соединяющий лобную и затылочную доли; • нижний продольный пучок, соединяющий затылочную и височную доли; • дугообразный пучок, соединяющий лобную долю и затылочно-височную кору; • крючковидный пучок, соединяющий лобную и верхнюю височную долю; • пояс, расположенный под корой поясной извилины.
1. Мозолистое тело. Мозолистое тело представляет собой самую крупную комиссуральную структуру, связывающую соответствующие участки правого и левого полушарий. Часть волокон идет из ствола мозолистого тела латерально и вверх, пересекая лучистый венец. Другая часть волокон проходит латерально и, изгибаясь, спускается вниз в виде тонкой пластинки волокон — покрова — к нижним участкам височной и затылочной долей. Парные волокна, идущие от валика мозолистого тела к медиальной стенке затылочной доли, образуют затылочные (большие) щипцы. Волокна, идущие от обеих сторон колена мозолистого тела к медиальной стенке лобной доли, называют лобными (малыми).
2. Малые комиссуральные структуры мозга. Передняя спайка связывает передние части височных долей с двумя обонятельными путями. Задняя и поводковая комиссуры располагаются непосредственно перед эпифизом. Спайка свода состоит из волокон, проходящих в ножках свода и соединяющих правый и левый гиппокампы.
Изображение коронарного среза мозга через переднюю ножку внутренней капсулы.Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше.Коронарный срез фиксированного препарата мозга. Уровень среза указан в верхней части изображения.Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше.
е) Боковые и III желудочки. Боковой (латеральный) желудочек состоит из тела (центральной части), расположенного в пределах теменной доли, и переднего (лобного), заднего (затылочного) и нижнего (височного) рогов. Передняя граница центральной части желудочка представлена межжелудочковым отверстием, расположенным между таламусом и передним столбом свода мозга. Посредством этого отверстия происходит сообщение III и латерального желудочков. Связь центральной части желудочка с задним и нижним рогами осуществляется через преддверие бокового желудочка.
Ниже перечислены связи, в которых участвует боковой желудочек.
• Передний рог. Границы рога формируют головка хвостатого ядра, прозрачная перегородка и структуры мозолистого тела (ствол мозолистого тела образует верхнюю стенку, колено мозолистого тела— переднюю стенку, клюв — нижнюю стенку).
• Тело. Тело бокового желудочка располагается ниже ствола мозолистого тела и выше таламуса и передней части тела свода. Медиально расположена прозрачная перегородка, сужающаяся в месте соединения мозолистого тела и приподнятой части свода. Прозрачная перегородка представляет собой истонченные стенки полушарий мозга. Наличие центральной полости в прозрачной перегородке подтверждает тот факт, что она состоит из двух частей.
• Задний рог. Задний рог располагается ниже валика мозолистого тела и медиальнее покрова мозолистого тела. На медиальной поверхности большие затылочные щипцы образуют луковицу заднего рога.
• Нижний рог. Нижний рог расположен снизу от хвоста хвостатого ядра и в передней части, ниже миндалевидного тела («миндалины»), входящего в состав лимбической системы. Дно рога формируют гиппокамп и соседние структуры.
• На внешней поверхности располагается коллатеральное возвышение, представляющее собой вдавление коллатеральной борозды в нижний рог.
III желудочек — полость промежуточного мозга. Границы желудочка показаны на рисунке ниже. Верхняя стенка желудочка представлена сосудистой оболочкой, от которой отходит сосудистое сплетение. Сосудистая оболочка III желудочка сформирована двойным слоем мягкой мозговой оболочки, сращенным с эпендимным эпителием желудочка. Сверху III желудочек граничит со сводом мозга и мозолистым телом. Боковые границы желудочка образованы таламусом и гипоталамусом. Передняя стенка желудочка представлена передней спайкой, терминальной пластинкой и перекрестом зрительных нервов. В нижней стенке различают воронку гипоталамуса, серый бугор, сосцевидные тела и верхний край среднего мозга. Заднюю стенку образуют эпифиз и прилежащие спайки.
Часто у людей старше 20 лет происходит кальцификация эпифиза и, в некоторых случаях, поводковой комиссуры, в связи с чем эти структуры становятся заметны даже на обзорной рентгенограмме черепа. Иногда происходит смещение эпифиза в полости черепа опухолью, гематомой или другим объемным новообразованием.
Система желудочков, изображенная под наклоном. Показана последовательность расположения структур в теле и нижнем роге бокового желудочка. Примечание: миндалевидное тело, краевая полоска и хвост хвостатого тела образуют крышу нижнего рога, а гиппокамп образует его дно (сосудистое сплетение удалено для обеспечения обзора).Коронарный срез через тело и нижний рог бокового желудочка.Левое полушарие мозга «на просвете», (А) медиальный и (Б) латеральный виды. (В) Коронарный срез, демонстрирующий расположение коротких и длинных ассоциативных волокон.Горизонтальный срез через колено и валик мозолистого тела. Волокна, идущие из ствола мозга латерально, пересекают лучистый венец.Система желудочков (А) изолированно и (Б) в структуре мозга.Коронарный срез фиксированного препарата мозга. Уровень среза указан в верхней части изображения.Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше.
Редактор: Искандер Милевски. Дата публикации: 9.11.2018
Проекционные волокна головного мозга
Проекционные волокна
Восходящие пути разделяются на: 1) экстероцептивные, начинающиеся от кожи и специализированных органов чувств; 2) проприоцептивные, начинающиеся от рецепторов надкостницы, капсул, связок суставов, мышц; 3) интероцептивные, начинающиеся от рецепторов внутренних органов и стенок кровеносных сосудов. Последние излагаются в разделе «Вегетативный отдел нервной системы».
Необходимы некоторые разъяснения и об осязании. Осязание, т. е. кожное «ощущение» (механорецепция), в действительности складывается из: 1) осязания (тактильное чувство), которое в силу быстрой адаптации реагирует только на первые моменты соприкосновения; 2) чувства давления; 3) чувства перемещения. Вторым обобщенным аналогичным понятием ощущений следует различать в системе экстерорецепции терморецепцию, имеющую специфические рецепторы чувства холода и тепла. У некоторых животных, например у гадюки, существуют специальные локальные рецепторы в области головы для принятия инфракрасного излучения. У человека тепловые рецепторы разбросаны по всей поверхности кожи.
Важным ощущением является ноцицепция, или болевое ощущение, возникающее при раздражении нервных окончаний в таламусе.
I. Экстероцептивные пути
1. Спиноталамический путь разделяется на латеральный и передний (tr. spinothalamici lateralis et anterior) (рис. 496). Содержит волокна, проводящие импульсы от кожных рецепторов, которые воспринимают тактильную, температурную и болевую чувствительность всех частей тела, кроме кожи и органов лицевого отдела головы. От кожи лица подобные раздражения воспринимаются рецепторами тройничного нерва. Первый нейрон спиноталамического пути униполярной формы, располагается в межпозвоночном узле. Его дендриты имеют свободные нервные окончания (воспринимают боль), тельца Краузе (воспринимают холод), тельца Руффини (воспринимают тепло). Аксоны в составе задних чувствительных корешков достигают клеток ядер заднего столба спинного мозга. Аксоны нейронов задних столбов через переднюю спайку проникают на противоположную сторону спинного мозга, поднимаясь на 2-3 сегмента, затем образуют пучок в передней части бокового канатика (передний спиноталамический путь), имея восходящее направление. В продолговатом мозге пучки волокон располагаются дорсальнее оливы, затем проникают в дорсальную часть моста, располагаясь рядом с медиальной петлей кнаружи от красных ядер. Этот нейрон заканчивается синапсом в переднелатеральном ядре таламуса, вступая в синаптическую связь с клетками III нейрона таламокортикального пути (tr. thalamocorticalis). Их отростки проникают через среднюю часть задней ножки внутренней капсулы, направляясь к задней центральной извилине (поля 1-2-3) и верхней теменной дольке (поля 5-7).
Центральные отростки I нейрона от клеток спинномозгового узла образуют латеральную группу волокон (латеральный спиноталамический путь), вступающих в спинной мозг. От места вхождения заднего корешка латеральная группа волокон имеет восходящие и нисходящие ветви на 1-2 сегмента (рис. 496). Волокна от ядер задних столбов спинного мозга переходят через переднюю спайку на противоположную сторону в передний канатик белого вещества; только в среднем мозге они присоединяются к волокнам латерального спиноталамического пути, достигая с ним таламуса и постцентральной извилины (корковый центр).
Вторыми нейронами пути болевой и температурной чувствительности являются клетки указанного ядра. Они совершают перекрест в мосту, продолговатом и спинном мозге, проходят в ретикулярной формации продолговатого мозга и моста, где, вероятно, имеют связи с ядрами, затем на противоположной стороне вступают также в состав медиальной петли, вместе с которой заканчиваются в переднем ядре таламуса. Третий нейрон этого пути располагается совместно с волокнами таламокортикального пути и достигает задней центральной извилины.
3. Проводящие пути обонятельного анализатора (tr. olfactorius) (рис. 497) имеют сложное строение и излагаются несколько схематизированно. Обонятельные рецепторы слизистой оболочки носовой полости воспринимают изменения химизма воздушной среды и являются самыми чувствительными по сравнению с рецепторами других органов чувств.
Корковый зрительный центр имеет весьма сложную многогранную систему нейронных связей. В ней находятся нейроны, реагирующие только на начало и конец освещения. В зрительном центре совершается не только обработка информации по ограничительным линиям, яркостям и градациям цвета, но и оценка направления движений объекта. В соответствии с этим и число клеток в коре головного мозга больше в 10000 раз, чем в сетчатке. Существенная разница имеется между числом клеточных элементов наружного коленчатого тела и зрительным центром. Один нейрон наружного коленчатого тела соединен с 1000 нейронов зрительного коркового центра, а каждый из этих нейронов в свою очередь образует синаптические контакты с 1000 соседних нейронов.
Аксоны II нейронов от клеток дорсального ядра образуют мозго-вые полоски (striae medullares ventriculi quarti), находящиеся в ромбовидной ямке на границе моста и продолговатого мозга. Большая часть мозговой полоски переходит на противоположную сторону и около средней линии погружается в вещество мозга, подключаясь к латеральной петле (lemniscus lateralis); меньшая часть мозговой полоски присоединяется к латеральной петле своей же стороны.
Аксоны II нейронов от клеток вентрального ядра участвуют в образовании трапециевидного тела (corpus trapezoideum). Большая часть аксонов переходит на противоположную сторону, переключаясь в верхней оливе и ядрах трапециевидного тела. Другая, меньшая, часть волокон оканчивается на своей же стороне. Аксоны ядер верхней оливы и трапециевидного тела (III нейрон) участвуют в образовании латеральной петли, в которой имеются волокна II и III нейронов. Часть волокон II нейрона прерывается в ядре латеральной петли (nucl. lemnisci proprius lateralis). Волокна II нейрона латеральной петли переключаются на III нейрон в медиальном коленчатом теле (corpus geniculatum mediale). Волокна III нейрона латеральной петли, пройдя мимо медиального коленчатого тела, заканчиваются в нижнем двухолмии, где формируется tr. tectospinalis. Те волокна латеральной петли, которые относятся к нейронам верхней оливы, из моста проникают в верхние ножки мозжечка и затем достигают его ядер, а другая часть аксонов верхней оливы направляется к мотонейронам спинного мозга и далее к поперечнополосатым мышцам.
6. Проводящий путь статокинетического аппарата (tr. vestibularis) (рис. 501). Осуществляет передачу импульсов при изменении положения головы и тела, участвуя совместно с Другими анализаторами в ориентировочных реакциях организма относительно окружающего пространства.
Первый нейрон статокинетическо-го аппарата находится в gangl. vestibulare, залегающем на дне внутреннего слухового прохода. Дендриты биполярных клеток преддверного узла формируют преддверный нерв, образованный 6 ветвями: rr. ampullaris superior, ampullaris lateralis, ampullaris inferior, ampullaris posterior, utricularis, saccularis; они контактируют с чувствительными клетками слуховых пятен и гребешков, расположенных в ампулах полукружных каналов, в мешочке и маточке преддверия перепончатого лабиринта.
Чувствительные клетки слуховых пятен и гребешков воспринимают смещение эндолимфы полукружных каналов и преддверия перепончатого лабиринта при малейшем изменении головы, при прямолинейном ускорении и вращении в трех плоскостях. Аксоны, т. е. центральные отростки биполярных клеток преддверного узла, образуют вестибулярную часть VIII пары черепных нервов совместно с n. cochlearis, которая покидает пирамиду височной кости через рorus acusticus internus, в мостомозжечковом углу проникает в вещество дорсальной части моста и продолговатого мозга, достигая верхнего, латерального, медиального и спинального ядер. Незначительное число волокон нейрона, минуя ядра, направляется непосредственно в мозжечок к nodulus, flocculus, uvula, culmen, lingula и nucl. fastigii. Между четырьмя вестибулярными ядрами существуют связи, а также двусторонние связи с ядрами ретикулярной формации.
Аксоны верхнего, латерального, медиального и спинального ядер преддверного нерва, помимо всех описанных нервных связей, образуют внутренние дуговые волокна в продолговатом мозге (fibrae arcuatae internae) и, подсоединившись к пучку медиальной петли, достигают латерального ядра таламуса, где и образуют синаптические контакты с III нейроном. От таламуса волокна направляются в корковые центры равновесия, находящиеся в средней височной извилине, лобной и теменной долях. Вероятнее всего, эти клетки рассеяны по всей коре головного мозга.
В таламусе располагаются III нейроны, аксоны которых проходят через заднюю ножку внутренней капсулы и оканчиваются в коре поля 43, в гиппокампе и крючке.
II. Проприоцептивные пути
1. Проводящие пути проприоцеп-тивной (глубокой) чувствительности. Состоят из пучков Голля и Бурдаха (рис. 502). С помощью этих путей совершаются движения, которые оцениваются сознанием. Управляемость движений осуществляется за счет афферентных импульсов из мышц и суставов движущихся частей тела. Импульсы достигают постцентральной извилины коры теменной доли. Эта обратная связь обеспечивает постепенность и координацию движений. При повреждении путей проприоцептивной чувствительности больной не может выполнять точные, соразмерные, ловкие движения.
Тонкий и клиновидный пучки оканчиваются не на ядрах спинного мозга, а в тонком и клиновидных ядрах продолговатого мозга. Аксоны клеток тонкого и клиновидного ядер (II нейрон) на границе с мостом образуют медиальную петлю, которая контактирует с клетками вентролатерального ядра таламуса. С латеральной стороны к медиальной петле присоединяются волокна спиноталамического пути. Аксоны из ядер таламуса (III нейрон), пройдя через заднюю часть внутренней капсулы, оканчиваются в коре верхней теменной дольки (поля 5 и 7) и в передней центральной извилине (поля 4-6).
Часть волокон II нейронов проприоцептивных чувствительных путей направляется в мозжечок через его нижние ножки, участвуя в механизме координации движений.
Существуют проприоцептивные чувствительные пути, которые связывают ядра спинного мозга, продолговатого мозга, моста, подкорковые образования, экстрапирамидную подсистему с мозжечком, участвующие в механизмах автоматической координации движений и тонусе мышц, помимо путей, замыкающихся в коре головного мозга. Эти механизмы, как правило, проявляются при внезапных нарушениях равновесия или выполнении автоматических движений (ходьба, танцы, письмо и др.), вырабатывающихся в процессе упражнений и под влиянием социальных моментов. Безусловнорефлекторные импульсы от всех перечисленных выше образований интегрируются в мозжечке, который координирует и определяет различные по точности движения. Импульсы из мозжечка оказывают регулирующее тормозящее влияние на ядра вестибулярного анализатора и ретикулярной формации. Так как от вестибулярных ядер возникает преддверно-спинальный путь, то по нему и ретикуло-спинальному пути наступает угнетение или облегчение функции альфа- и гамма-мотонейронов передних столбов спинного мозга и мышечных веретен двигательных периферических нервов. Таким образом, благодаря механизмам обратной связи через вестибуло-спинальный и ретикулоспинальный пути мозжечок согласовывает быстрые и медленные сокращения всех мышц. Мозжечок напоминает блок регуляции, основанный на принципе обратной связи. Червь мозжечка координирует движения при ходьбе и стоянии. В полушарии мозжечка расположены механизмы очень точной координации движений, преимущественно для выполнения движений верхней конечностью. Червь подчинен коре мозжечка, а она функционирует под влиянием коры головного мозга.
2. Проводящие пути от спинного мозга к мозжечку. Задний спинно-мозжечковый путь, или путь Флексига (tr. spinocerebellaris dorsalis) (рис. 503). Имеет веретеновидные рецепторы в скелетных мышцах. Его первый специализированный чувствительный нейрон располагается в спинномозговом узле. Аксоны в составе заднего корешка достигают клеток Кларка основания заднего рога, расположенных на уровне от VII шейного до II поясничного сегмента спинного мозга, где вступают в синаптическую связь со II нейронами. Аксоны II нейронов образуют на своей стороне пучок волокон, расположенный в задней части бокового канатика (латеральнее пирамидного пути), через нижние ножки мозжечка вступают в синаптическую связь с клетками коры червя мозжечка. Из коры червя начинается III нейрон, оканчивающийся у клеток зубчатого ядра мозжечка.
3. Передний спинно-мозжечко-вый путь, или путь Говерса (tr. spinocerebellaris ventralis) (рис. 503). Его рецепторы также начинаются в мышцах. Первый специализирован-ный чувствительный нейрон располагается в спинномозговых узлах. После переключения на II нейроны в промежуточно-медиальном ядре спинного мозга их нервные волокна через переднюю серую спайку спинного мозга переходят на противоположную сторону и располагаются в передней части бокового канатика. В мосту они находятся в его дорсальном отделе, медиальнее лицевого нерва. В среднем мозге спинно-мозжечковый вентральный путь достигает нижнего двухолмия и направляется через верхние ножки мозжечка своей же стороны к коре червя. Из червя начинается III нейрон, заканчивающийся в зубчатом ядре мозжечка. Таким образом, при одностороннем включении заднего и переднего спинно-мозжечковых путей расстройства координации возникают на обеих сторонах тела.
4. Проводящие пути от мозгового ствола к мозжечку. Преддверно-мозжечковый путь (tr. vestibulocerebellaris). Начинается преимущественно от ядра Дейтерса (nucl. lateralis) и в составе нижних ножек мозжечка достигает шатрового ядра (n. fastigii) червя мозжечка.
5. Ретикуломозжечковый путь (tr. reticulocerebellaris). Аксоны идут из ядер ретикулярной формации продолговатого мозга через нижние ножки мозжечка к его ядрам, а от ретикулярной формации моста проходят через средние ножки мозжечка к его ядрам.
6. Оливомозжечковый путь (tr. olivocerebellaris). Начинается от оливы продолговатого мозга, после частичного перекреста направляется через нижние ножки мозжечка и оканчивается в его коре.
7. Путь от ядер задних столбов продолговатого мозга к мозжечку. От нежного и клиновидного ядер (nucll. gracilis et cuneatus) продолговатого мозга начинаются волокна, которые формируют наружные дуговые волокна (librae arcuatae externае), неперекрещенные, достигающие через нижние ножки мозжечка коры червя.
Проводящие пути от коры головного мозга к мозжечку.
8. Лобно-мостовой путь (tr. frontopontinus). Начинается в лобной доле. Его волокна погружаются в белое вещество полушарий, подключаясь к системе волокон corona radiata, затем располагаются в передней ножке внутренней капсулы, внутреннего отдела основания ножки мозга, заканчиваясь в ядрах моста своей же стороны.
а) Затылочно-височно-мостовые пути (trr. occipitotemporopontini). Формируются в коре одноименных долей, направляясь в глубину полушарий, где проходят через задние ножки внутренних капсул. В среднем мозге располагаются в латеральном отделе ножек мозга, затем переключаются в ядрах моста своей же стороны.
б) Мостомозжечковые пути (tr. pontinocerebellaris). Начинаются от ядер моста, аксоны которых перекрещиваются и, соединившись в средних ножках мозжечка, достигают коры его полушарий. Поэтому расстройства координации, возникающие при одностороннем поражении лобных и височно-затылочных долей коры головного мозга, выявляются на противоположной стороне очага поражения.
9. Проводящие пути от мозжечка. Мозжечково-красноядерный путь (tr. cerebellorubralis). Аксоны клеток коры мозжечка переключаются в зубчатом ядре. Одна часть волокон через верхние ножки мозжечка после перекреста в среднем мозге достигает красных ядер. Другая часть волокон направляется в таламус. Из таламуса импульсы достигают задней центральной извилины коры. За счет этих волокон осуществляется информация о выполненной жорректировке автоматических импульсов от красного ядра, достигающих спинного мозга по красноядерно-спинномозговому пути.
10. Нисходящие (проекционные) двигательные проводящие пути, идущие от коры головного мозга к спинному и двигательным ядрам черепных нервов, разделяются на пирамидные, экстрапирамидные и волокна, проводящие торможение.
а) Пирамидный путь (tr. pyramidalis) (рис. 504). Хорошо развит у человека, так как по нему передаются импульсы поперечнополосатым мышцам при выполнении целенаправленных, тонко координированных сознательных движений. Пирамидные пути существуют и у многих животных, но функционируют без сознательной корректировки. Двигательные клетки коры не иннервируют отдельно ту или другую мышцу, а осуществляют заданную программу движений для отдельных групп мышц. Пирамидный путь получил свое название от двух клиновидной формы выпуклостей, лежащих на вентральной поверхности продолговатого мозга. Многие годы считалось, что все волокна пирамидного пути начинаются от клеток коры передней центральной извилины. В настоящее время установлено, что только около 40% аксонов, проходящих через пирамиды, начинается от клеток двигательной зоны коры, а 20% аксонов пирамидного пути начинается от клеток задней центральной извилины (соматосенсорная область). Остальные 40% волокон присоединяются к пирамидному пути от клеток различных областей коры полушарий большого мозга.
Первые нейроны располагаются в передней центральной извилине, предцентральной и парацентральной дольках (поля 4-6), часть нейронов разбросана в других корковых полях (7-8-9-22-24 и др.). Существенным моментом является то, что все корковые поля пирамидного пути связаны с нейронами, которые своей деятельностью подавляют моторную активность двигательной зоны и находятся в полях 2-4-8-19. Подобная тормозящая система отсутствует у других проводящих путей. Кроме того, в поле 4 есть участок 4S, откуда специальные аксоны достигают ядер ретикулярной формации, которая оказывает тормозящее или возбуждающее влияние на произвольные рефлексы. Дендриты пирамидных клеток имеют связь со вставочными нейронами, подключающими чувствительные клетки всех анализаторов. Эти вставочные нейроны формируют короткие и длинные ассоциативные пути белого вещества.
Пирамидный путь включает три пучка: а) корково-ядерный путь (tr. corticonuclearis), осуществляющий центральное кодирование программы движений в двигательных ядрах черепных нервов (III, IV, V, VI, VII, IX, X, XI, XII пары); б) передний корково-спинальный путь (tr. corticospinalis anterior); в) боковой корково-спинальный путь (tr. corticospinalis lateralis). Оба последних пучка проводят импульсы программы движений к мотонейронам спинного мозга.
Первые нейроны пирамидного пути находятся в различных областях коры полушарий головного мозга. В V слое коры головного мозга имеются пирамидные клетки Беца, аксоны которых принимают участие в образовании лучистого венца белого вещества полушарий мозга. Эти волок-на конвергируют вниз, проходя в колене и в 2/3 задней ножки внутренней капсулы. Пирамидные клетки имеют длинные аксоны и большое число коллатералей, которые подключают несколько двигательных клеток II нейронов.
Волокна пирамидного пути, пройдя внутреннюю капсулу, располагаются в основании ножки мозга, где от них отделяются перекрещенные волокна к ядрам глазодвигательного нерва (иннервирующего, верхнюю, нижнюю, медиальную прямые, нижнюю косую мышцы глазного яблока и мышцу, поднимающую верхнее веко), к ядру блокового нерва (иннервирующего верхнюю косую мышцу глазного яблока) и к ядру отводящего нерва (иннервирующего латеральную прямую мышцу глазного яблока).
Из основания ножки мозга пирамидный путь спускается в вентральную часть моста, на уровне которого отделяются перекрещенные волокна для контакта с двигательным ядром тройничного нерва (иннервирующего жевательные мышцы), с двигательным ядром лицевого нерва (иннервирующего мимическую мускулатуру); некоторые волокна отдают коллатерали в ретикулярную формацию. Пучок пирамидного пути расположен в мосту некомпактно, через него поперечно проходят волокна корково-мостомозжечкового пути (описан в разделе «Проприоцептивные пути»). В продолговатом мозге волокна пирамидного пути объединяются в компактный пучок и на вентральной поверхности продолговатого мозга образуют пирамиды. В каждом из двух трактов пирамидных путей содержится около 1 млн. волокон, главным образом тонких и скудно миелинизированных; около 3% волокон имеет большой диаметр и покрыто толстой миелиновой оболочкой; они являются аксонами клеток Беца. В продолговатом мозге двигательные ядра языкоглоточного (IX пара), блуждающего (X пара), добавочного (XI пара), подъязычного (XII пара) нервов также контактируют с волокнами пирамидного пути. Волокна пирамидного пути, направляющиеся к ядрам двигательных черепных нервов, перекрещиваются. Эти ядра получают иннервацию от волокон своей и противоположной сторон. Поэтому при центральном одностороннем поражении коры полушарий мозга или проводящих путей не бывает полного паралича мышц, иннервируемых III, IV, V, VI, VII, IХ, X, XI парами черепных нервов. В области пирамид продолговатого мозга небольшая часть волокон пирамидного пути, огибая нижнюю оливу через нижнюю или среднюю ножку мозжечка, входит в него.
Следовательно, пирамидный путь осуществляет главным образом перекрестную иннервацию. Поражение бокового корково-спинномозгового пути вызывает расстройство движений конечностей на противоположной стороне и почти не нарушает функции мышц туловища вследствие сохранения иннервации за счет переднего кортико-спинального пучка. Такой односторонней иннервацией обладают не все мышечные группы. Большая часть мышц, а именно мышцы глазного яблока, жевательные, мимические мышцы верхней части лица, глотки, гортани, шеи, туловища и промежности, имеет двустороннюю иннервацию за счет волокон перекрестных и своей стороны. Односторонне иннервируются мышцы конечностей, языка, мимические мышцы ниже ротовой щели. Поражение соответствующих клеток коры вызывает полный паралич.
б) Экстрапирамидные пути. Подразделение двигательной системы на пирамидную и экстрапирамидную является источником путаницы и ошибок, так как вначале пирамидную систему представляли единственной двигательной системой. Вероятно, поэтому все двигательные пути, описанные позднее, были объединены в экстрапирамидную систему. Провести четкую грань в анатомическом строении и функциональном значении между пирамидной и экстрапирамидной системами практически невозможно, так как они функционируют в полном взаимодействии. С помощью экстрапирамидной системы тело поддерживается в нужном положении и выполняются содружественные движения с регуляцией мышечного тонуса при выполнении безусловнорефлекторных двигательных актов.
К экстрапирамидной системе относятся многие рассеянные двигательные клетки среди чувствительной зоны, базальные ядра (бледный шар, скорлупа, хвостатое ядро, ограда), передненижнее ядро таламуса, субталамические ядра, мозжечок, красное ядро, черная субстанция и ядра ретикулярной формации.
Экстрапирамидная проводящая система начинается от разбросанных по всей коре головного мозга двигательных клеток, аксоны которых поступают через заднюю ножку внутренней капсулы к хвостатому ядру и скорлупе, а затем после переключения идут к бледному шару. От наружного членика бледного шара аксоны направляются в черное вещество, красное ядро, медиальное вестибулярное ядро и ретикулярную формацию.
От ядер ретикулярной формации берет начало ретикулярно-спинно-мозговой путь (tr. reticulospinalis), вступающий в контакт со вставочными нейронами и малыми альфа-нейронами передних столбов спинного мозга. От красных ядер формируется tr. rubrospinalis, входящий в боковой канатик спинного мозга и контактирующий со вставочными нейронами заднего и переднего столбов спинного мозга, которые переключаются на малые альфа- и гамма-нейроны.
В экстрапирамидной системе имеется замкнутая петля проводящих путей; она начинается в бледном шаре и проходит к таламусу, где есть центр «активации», затем к скорлупе и вновь возвращается к бледному шару. Бледный шар связан с моторными клетками коры, которые и активируются под действием возвратных импульсов, посылая прямые сигналы к спинному мозгу через пирамидные пути. Допускается, что полосатые тела (хвостатое ядро, скорлупа, бледный шар) являются первичными инициаторами двигательной активности после получения сигналов из сенсорных областей. Кора не доминирует над полосатым телом, но в процессе формирования движений повышает способность экстрапирамидной системы к включению в рефлекторный процесс и увеличивает ее гибкость в процессе перестройки.
в) Волокна, проводящие торможение разделяются на центральные и периферические.
Центральное торможение формируется специализированными нейронами ядер стволовой части мозга и коры полушарий. При централь-ном торможении импульсы идут от высших центров по эфферентным нервным волокнам к сенсорным органам. Среди избытка информации тормозные импульсы играют большую роль в отборе воспринимаемых импульсов головным мозгом. В этом случае при переключении внимания на какой-нибудь стимул импульсы других сенсорных систем угнетаются импульсами торможения, направляя необходимые стимулы в определенный «канал». За счет торможения производится фильтрация импульсов и их суммация, что обеспечивает локальность возбуждения и его целенаправленность. Импульсы торможения создают гиперполяризацию рецепторной мембраны, тем самым инактивируют рецептор, что нарушает восприятие раздражений.
Периферическое торможение возникает при взаимодействии сенсорных клеток, действуя на сами сенсорные клетки или синапсы. Периферическое торможение осуществляется за счет латеральных нервных волокон, соединяющих соседние рецепторы и клетки. Например, при нагрузке на мышцу сокращение развивается равномерно во всех экстрафузальных мышечных волокнах. Это распределение осуществляется за счет латеральных нервных волокон, при помощи которых возбужденный мотонейрон тормозит себя и другие нейроны соответствующего двигательного пула, возбуждая тормозные клетки Реншоу. Аксоны клеток Реншоу в свою очередь также заканчиваются на мотонейронах. Петля обратной связи тормозит повторное быстрое возбуждение мотонейронов передних столбов спинного мозга.
В действительности иннервация поперечнополосатых мышц значительно сложнее, чем это можно изложить при описании проводящих путей.
Каждая мышца человека одинаково хорошо снабжена чувствительными и двигательными нервами. Один мотонейрон спинного мозга иннервирует одновременно большое число мышечных волокон, объединяя их в двигательную единицу. Естественно, что в иннервации мышц принимают участие несколько двигательных единиц. Мотонейроны, обеспечивающие иннервацией одну мышцу, не представлены единым компактным ядром, а рассеяны по нескольким сегментам спинного мозга и функционально объединяются в двигательный пул. В пределах каждого двигательного пула имеется сложная система вставочных нейронов, объединяющая в одно целое альфа-и гамма-мотонейроны спинного мозга. Этот комплекс вставочных нейронов частично описан при изложении бокового кортико-спинального пути и рефлекторной дуги.
Чтобы понять принцип иннервации мышц, необходимо кратко познакомиться со строением мышечной ткани. Каждая поперечнополосатая мышца имеет неоднородные экстрафузальные мышечные волокна, иннервируемые концевыми пластинками двигательных нервов. Кроме того, в меньшем числе встречаются тонкие интрафузальные мышечные волокна, образующие мышечные веретена. В каждом мышечном веретене может быть по 2-10 интрафузальных мышечных волокон, лежащих в концевых частях веретена. Задачей интрафузальных мышечных волокон является натяжение мышечной трубки, где находится первичный чувствительный рецептор. Эта трубка расположена между двумя сократительными концами веретена (в них находятся интрафузальные мышечные волокна) и чрезвычайно чувствительна к малейшему сокращению интрафузальных волокон или растяжению мышцы. Концы мышечных веретен прикреплены к оболочке экстрафузальных волокон, так что при малейшем изменении их длины меняется натяжение мышечных веретен и соответственно их мышечных трубок. При изотоническом сокращении мышечное волокно укорачивается и натяжение на первичном рецепторе уменьшается. Когда мышца расслабляется, ее веретена растягиваются и первичные рецепторы возбуждаются. Следовательно, первичный рецептор может растягиваться в 3 случаях: при расслаблении экстрафузальных волокон, сокращении интрафузальных волокон и растяжении мышц за счет внешней нагрузки. Аксон, идущий от первичного рецептора мышечного веретена, возвращается в соответствующий двигательный пул, из которого выходит нерв к экстрафузальным мышечным волокнам. При растяжении мышечной трубки в ней генерируются импульсы, которые поступают в задние столбы спинного мозга.
После короткой задержки возникает внезапное сокращение четырехглавой мышцы. При длительном изотоническом сокращении мышцы крупные мотонейроны спинного мозга должны постоянно генерировать импульсы для сокращения экстрафузальных мышечных волокон. В этом случае поступающие даже слабые импульсы из моторной зоны коры головного мозга возбуждают только мелкие мотонейроны, которые вызывают сокращение интрафузальных мышечных волокон. Оно приводит к натяжению мышечных трубок и генерации импульсов для возбуждения крупных мотонейронов всего двигательного пула спинного мозга данной мышцы. Таким образом автоматически поддерживается изотоническое сокращение.
Механизм обратной связи мышечного веретена является весьма эффективным в случаях запредельных сокращений. Теоретически допустимо, что вполне возможно повреждение сухожилия и даже костей при максимальном сокращении, когда возбуждаются достаточно сильно крупные мотонейроны двигательного пула спинного мозга. Подобных запредельных сокращений в здоровом организме не происходит, так как в сухожилиях мышц имеются рецепторы Гольджи. Эти рецепторы выдают тормозные залпы, вызывающие угнетение мотонейронов спинного мозга.
 Изображение коронарного среза мозга через переднюю ножку внутренней капсулы.
Изображение коронарного среза мозга через переднюю ножку внутренней капсулы. 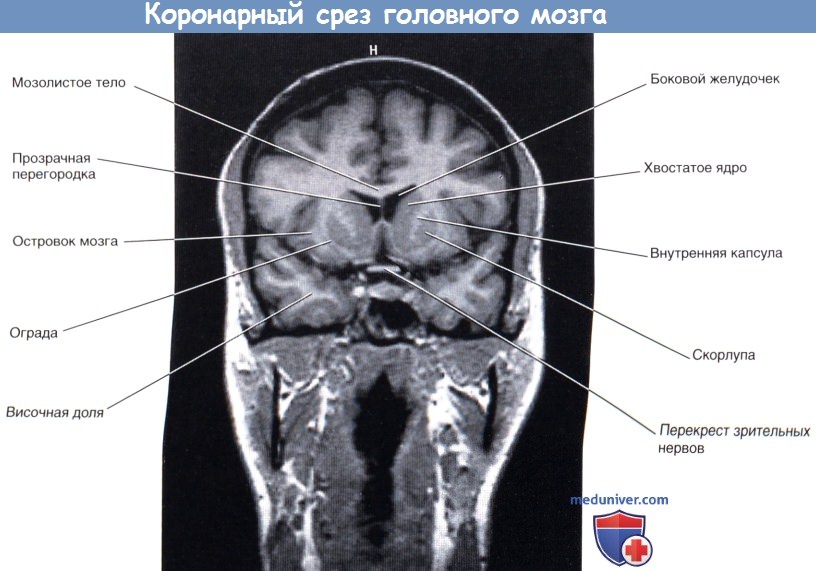 Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше.
Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше. 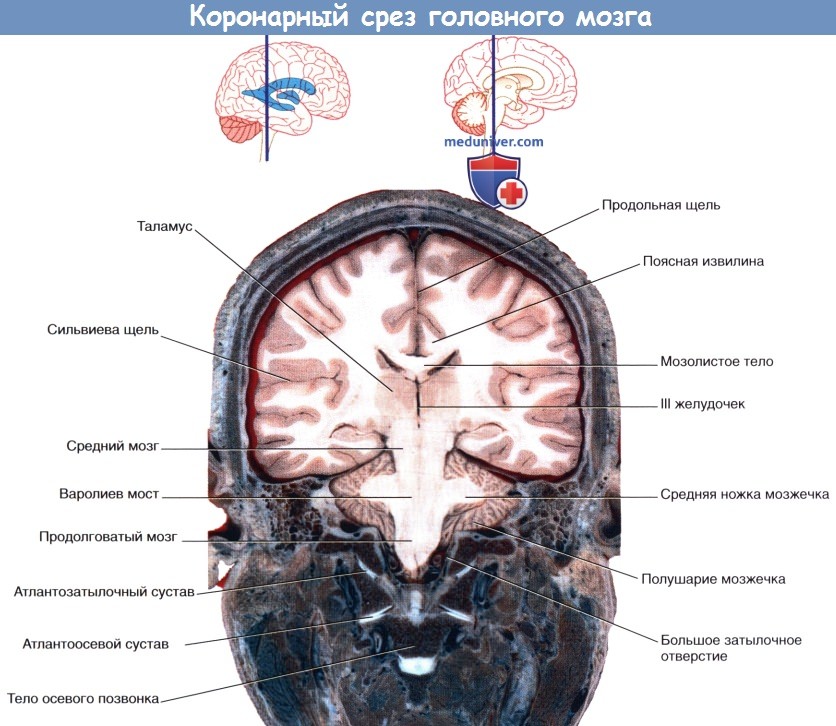 Коронарный срез фиксированного препарата мозга. Уровень среза указан в верхней части изображения.
Коронарный срез фиксированного препарата мозга. Уровень среза указан в верхней части изображения. 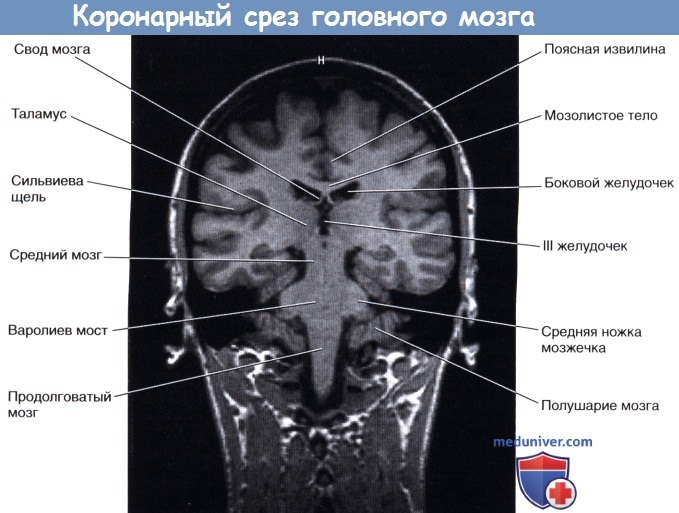 Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше.
Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше. 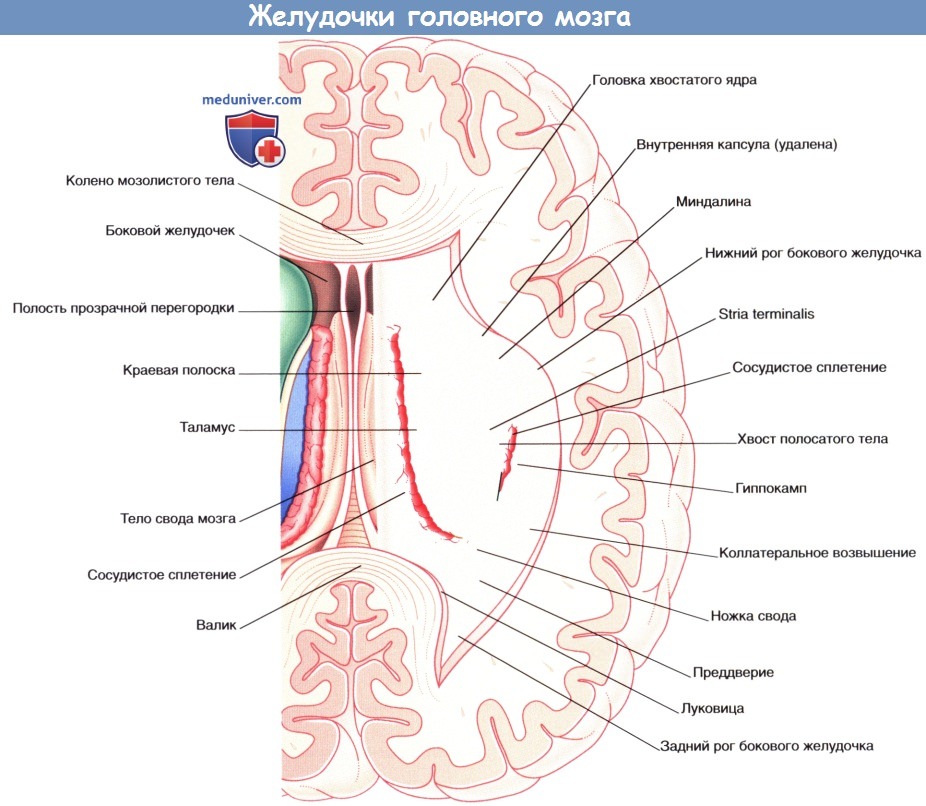 Система желудочков, изображенная под наклоном. Показана последовательность расположения структур в теле и нижнем роге бокового желудочка.
Система желудочков, изображенная под наклоном. Показана последовательность расположения структур в теле и нижнем роге бокового желудочка. 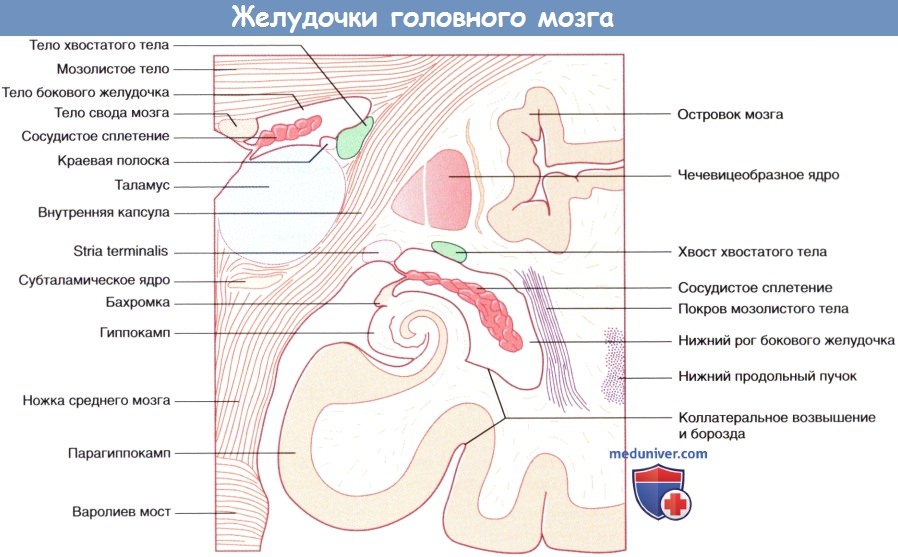 Коронарный срез через тело и нижний рог бокового желудочка.
Коронарный срез через тело и нижний рог бокового желудочка. 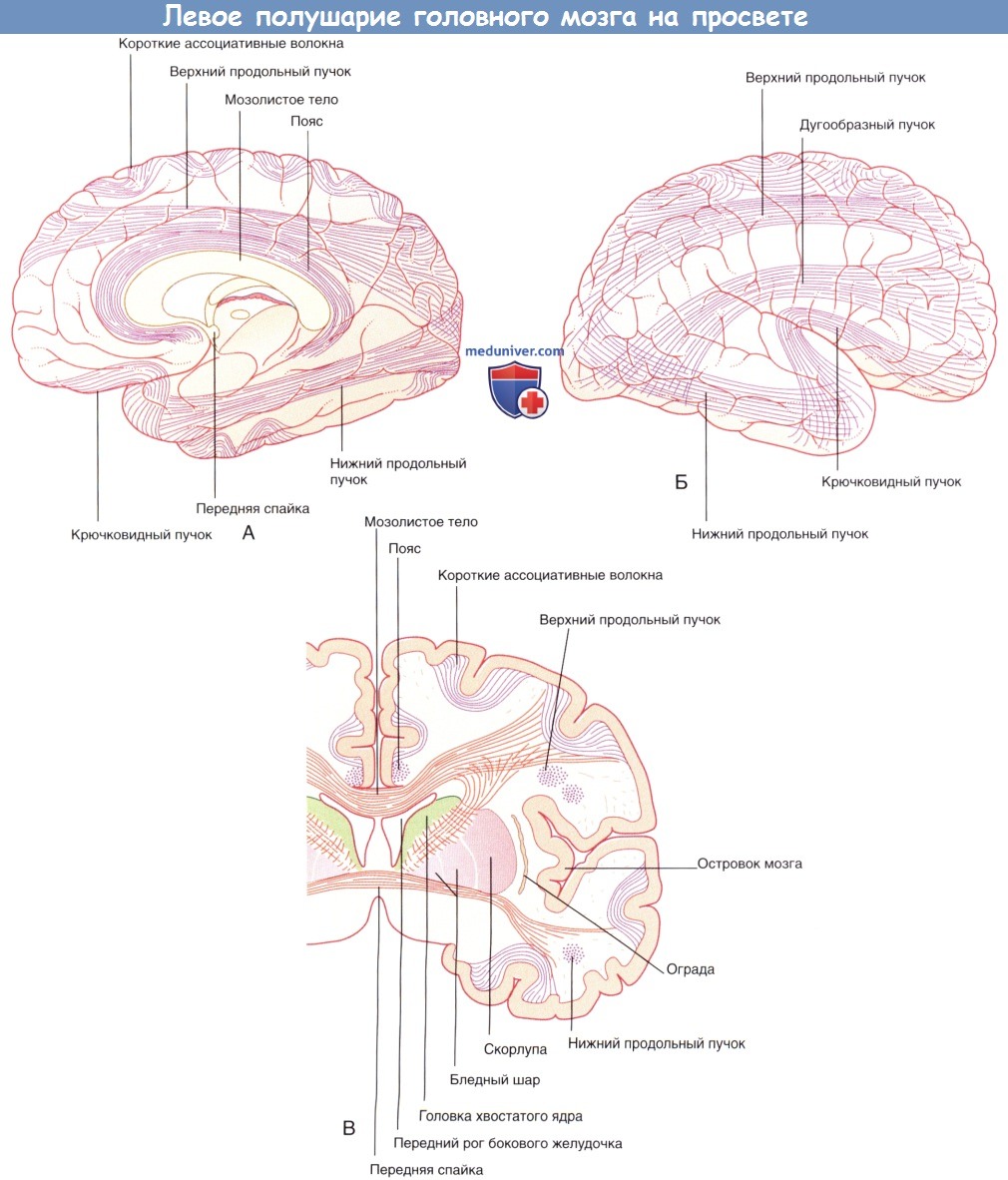 Левое полушарие мозга «на просвете», (А) медиальный и (Б) латеральный виды.
Левое полушарие мозга «на просвете», (А) медиальный и (Б) латеральный виды. 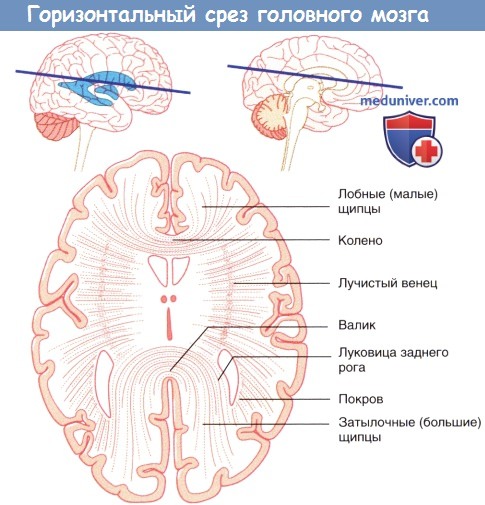 Горизонтальный срез через колено и валик мозолистого тела.
Горизонтальный срез через колено и валик мозолистого тела. 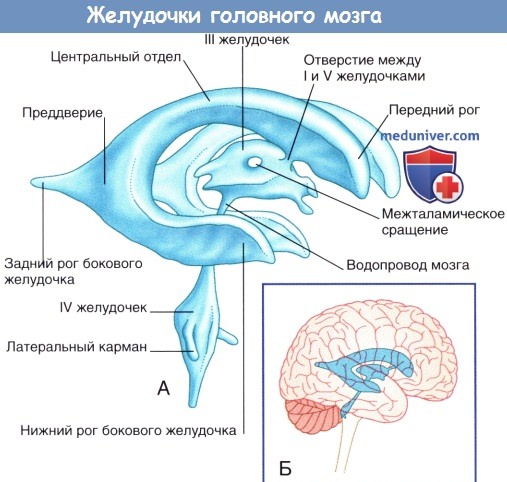 Система желудочков (А) изолированно и (Б) в структуре мозга.
Система желудочков (А) изолированно и (Б) в структуре мозга. 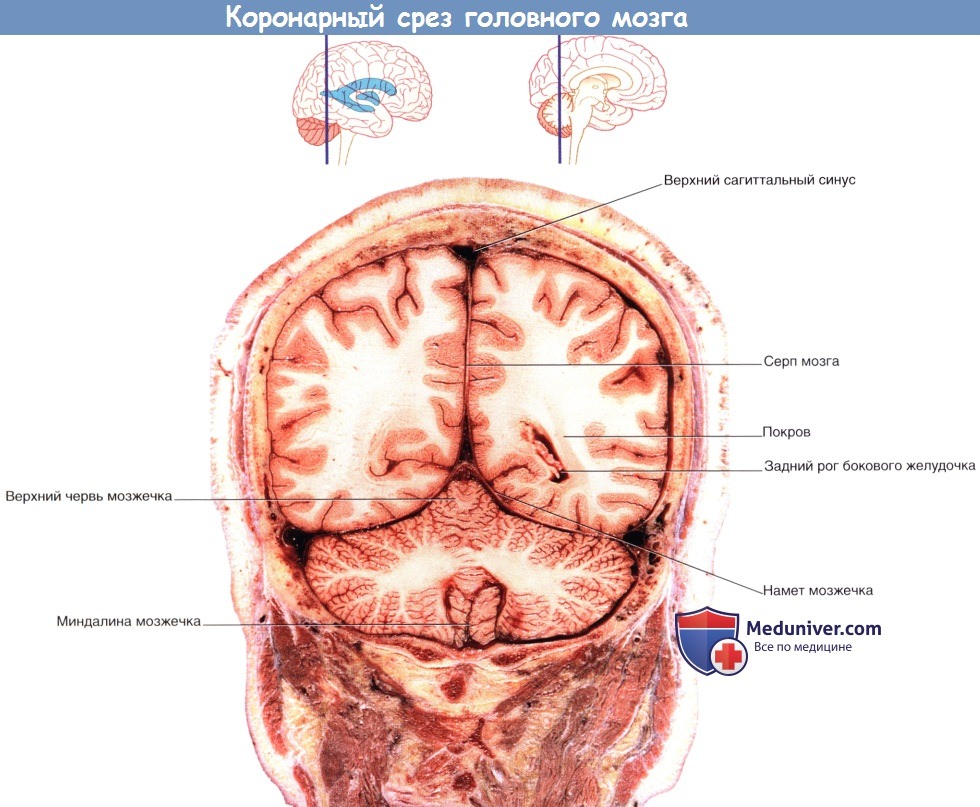 Коронарный срез фиксированного препарата мозга. Уровень среза указан в верхней части изображения.
Коронарный срез фиксированного препарата мозга. Уровень среза указан в верхней части изображения. 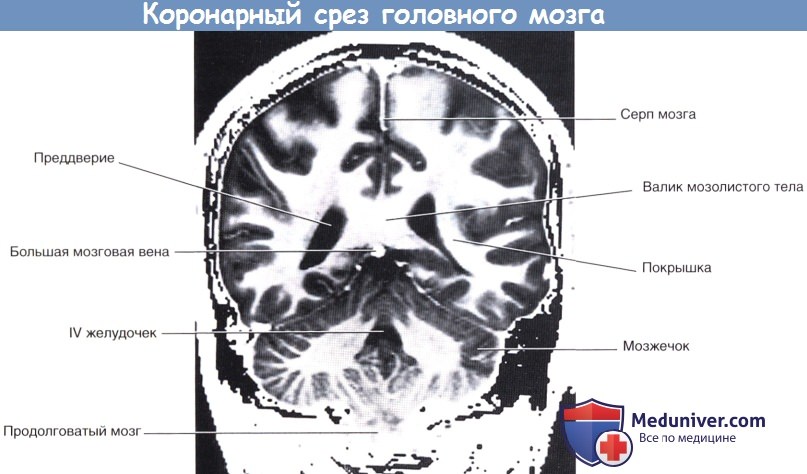 Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше.
Коронарный МР-срез в месте, аналогичном уровню среза рисунка выше. 


